



Cleavage of Microlecithal Eggs
This page shows a series of photographs which follows a microlecithal (oligolecithal) egg from the unfertilized single cell through to the late blastula. Microlecithal eggs have a very small amount of stored yolk, distributed evenly throughout the cytoplasm. As a result of this small amount of yolk, these animals show one of two different methods of development. The first is to have a feeding larval stage (as this embryo will have) to meet the nutritional requirements of the embryo, with the second being to have internal development such that the nutritional requirements of the embryo are met by the mother (what we will see later with mammals). These particular slides are whole mounts of starfish embryos.
Later
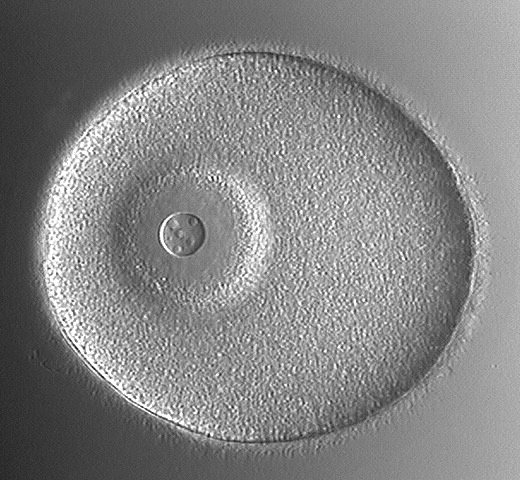
This picture is of the unfertilized egg. It can be differentiated from the zygote by the presence of a large, conspicuous nucleus (large arrow) with obvious nucleolus (smaller arrow) and by the lack of a fertilization membrane. This shows the zygote (fertilized cell). It is recognized by the presence of the fertilization membrane (arrows) surrounding it and the peripheral, fluid-filled perivitelline space.
Later
This is the two cell stage. By this stage, the zygote has completed its first cleavage, which is both equal and holoblastic (i.e. the entire ovum is divided into cells). The division (cleavage) has passed through the animal-vegetal axis, producing two similar blastomeres. This is the four cell stage. This second cleavage also passes through the animal-vegetal axis, but perpendicular to the first cleavage (described previously). Four equal-sized blastomeres are the end result.
Later
The eight cell stage. Here, the third cleavage has occurred in the equatorial plane (perpendicular to the first two cleavages and the animal-vegetal axis). Note that the upper four blastomeres (the animal pole) are slightly smaller than the lower four blastomeres (the vegetal pole). NOTE: The sixteen cell stage is not shown - its division is once again vertical, along the animal-vegetal axis. The thirty-two cell stage is seen here. After the 16 cell stage, the cleavages become more difficult to follow, due to the increasing number of cells and to the division of blastomeres becoming asynchronous. Cleavage continues, forming a mass of cells which organizes itself into the blastula. The lighter area in the centre of the embryo is the beginning of the blastocoel.
Later
This shows the early blastula. With continuing cleavage, the cells in centre begin to lose contact with one another, and a central fluid-filled cavity (the blastocoel) forms. This blastocoel is surrounded by a single layer of cells, forming the hollow sphere know as the blastula. This shows the late blastula. Like the early blastula, it is characterized by a single layer of cells surrounding the central hollow area - the blastocoel (B). The blastomeres are seen to be smaller and are individually not as obvious. The blastomeres at the vegetal pole (VP) are taller than those at the animal pole (AP), making the vegetal pole appear slightly thicker.
Gastrulation in the Microlecithal Embryo
On this page, a series of photographs can be seen which follow a microlecithal (oligolecithal) embryo from the early gastrula stage through gastrulation to the early bipinnaria larval stage.
The process of gastrulation is affected, as was cleavage, by the amount and the distribution of yolk. As a result, gastrulation differs between different species in details, but the principle is the same in all these species. During gastrulation, some of the cells from the surface of the embryo move to the interior and replicate, thereby forming new layers of cells. These movements are coupled with the differentiation of the migrating cells (caused by the differential activity of certain genes) into histologically unique layers.
During microlecithal gastrulation, a blastula with a relatively large blastocoel is usually formed. As a result, there is plenty of space to move some of the surface cells to the inside to form new layers. This is first seen as the starfish embryo becomes flattened at the vegetal pole (the initial migration and differentiation of cells), which will then invaginate within the blastocoel. When this occurs, the internal layer of cells formed (called the hypoblast or mesendoderm) are the cells which gives rise to the endoderm and mesoderm germ layers. The cells who remain on the outer surface of the gastrula become the epiblast (ectoderm). The cavity created by the invagination is called the gastrocoel or archenteron (primitive gut), whose opening at the vegetal pole is called the blastopore. The anterior end of the hypoblast (mesendoderm) develops out-pocketings (evaginations) on each side to form the mesoderm (mesodermal pouches). These pouches are also called enterocoelic pouches and the cavities within them are called the coelomic cavities. As the embryo grows during gastrulation, cells also bud off the hypoblast and migrate into the blastocoel to form mesenchymal cells.
Later
This is the early starfish gastrula. The vegetal pole (VP) is showing the first signs of invagination. AP=animal pole, VP=vegetal pole. This shows the early gastrula. The beginning of the invagination which will form the gastrocoel (a.k.a. archenteron) is obvious now, as it invaginates anteriorly (toward the animal pole (AP)) from the vegetal pole (VP), where the blastopore is formed.
Later
Here we see the mid gastrula. The gastrocoel continues to develop as the embryo grows. AP=animal pole, VP=vegetal pole. This is the late gastrula. This picture is a dorsal view of the embryo, with anterior being toward the animal pole (AP) and posterior toward the vegetal pole (VP). Note the beginning of the formation of enterocoelic pouches on the right and left side of the anterior end of the gastrocoel.
C
O
L
O
U
R
I
Z
E
!!
This is the early bipinnaria larva seen in lateral view (from the side). The dorsal aspect of the embryo is facing right and the ventral aspect is on the left side of this picture. The blastopore is at the posterior end (bottom) and is now developing into the anus, while the anterior end of the gastrocoel has bent toward the ventral aspect of the embryo, where a mouth has formed. Here we see the same picture as on the right, but now labelled and in glorious technicolour!
Blastocoel
Enterocoelic pouch
Mouth
Archenteron
Blastopore (anus) blue=Ectoderm
red=Mesoderm
yellow=Endoderm
Cleavage and Morphogenesis
Development of Sea Star Embryos (Rust, fig. 110)
Egg and Zygote
The unfertilized egg (1a) has a large and conspicuous nucleus. Compare the amount of yolk and cytoplasm to the size of the nucleus. The nucleus of the zygote (1b) is less conspicuous. The fertilization membrane is not present in this preserved material
.
cell stage
Successive cell divisions divide the zygote into smaller and more numerous cells. Compare the size of the 2,4 and 8 cell stages with the size of the original zygote. Note the cleavage furrows formed by the pinching in of the cell membranes during division. From the arrangement of the cells determine the plane of cleavage that produced the cells.
.
Early blastula
Cleavage continues producing many additional cells which arrange themselves around a fluid filled cavity (blastocoel). The blastula (7) begins to take on the appearance of a ball.
. Blastula
This ball-like stage is called the blastula (5). Compare the size of the blastula to the original zygote. Note the blastocoel (6).
late gastrula
During the late blastula stage a few cells on one side of the developing embryo begin to push into the blastocoel (8). When this process (gastrulation) becomes visible as an indentation, the developing embryo is termed a gastrula (9). The point of indentation is termed the blastopore (10). As the invagination process continues (11),with more cells pushing into the interior, a new cell layer is formed and a new cavity (12) (archenteron) appears within this new layer of cells. This cavity opens through the blastopore (14) to the outside of the embryo. Later a third layer of cells develops between the original two layers. These three cell layers (ectoderm, endoderm and mesoderm) provide the "bricks and mortar" for the differentiation of the embryo's organ systems.
. Larval
The developmental process to the end of the gastrula (13) stage takes approximately two days. At this time the gastrula transforms itself into a free swimming (via cilia bands) (18) and feeding larva (15). This change includes the formation of a digestive system with two openings (mouth (17) and anus (16)). The larva grows as it feeds on algae and diatoms. After several months the larva undergoes a process called metamorphosis that transforms it into a tiny sea star.
Sea stars belong to the Phylum Echinodermata, and display the typical deuterostome pattern of early development. In sea stars, fertilization and development take place externally and the eggs carry little yolk, so it is easy to observe early development. Obtain a slide of developing sea star embryos and
identify the following developmental stages.
_Cleavage
The earliest stages are simply named for the number of cells they contain: the 2-cell stage, the 4-cell stage, the 8-cell stage, and so on. These stages result from the first cleavage, the second cleavage, the third cleavage, etc. (Individual cells that form during these cleavages are called blastomeres (Gr: blastus = a bud, mere = part).) Once the cells become too numerous to be counted easily, which usually happens around the 32- or 64-cell stage, this terminology is abandoned and the embryo is said to be in the morula (Latin: little mulberry) stage. The morula is a spherical mass of from a few dozen to several hundred cells.
Blastulation
The morula is solid, but as cleavage continues the blastomeres move toward the periphery of the mass, leaving a fluid-filled cavity in the interior. Soon the embryo no longer resembles a simple mass of cells, but instead takes on the form of a sphere. In this stage the embryo is called a blastula (little bud), and the fluid-filled cavity is called a blastocoel (Gr: coelo = hollow). This change takes place about ten hours after fertilization. When the blastula first forms it contains about 1000 cells.
Gastrulation
A few hours after the formation of the blastula the process of gastrulation begins. At a certain point on the surface of the blastula cells begin to move toward the center of the blastocoel, so that an opening is formed leading into a blind or dead-end tube. If you were to take a rubber balloon loosely filled with water or air and poke your finger into it without breaking the balloon you would be at least roughly approximating the process of gastrulation. The opening on the surface of the embryo is called the blastopore, and in deuterostomes such as sea urchins and sea stars this will become the anus of the mature organism. The blind or dead-end tube is called the archenteron (Gr: archi = first, enterum = intestine). This will eventually reach all the way through and open out the other side, and thus will form the basis of the animal's digestive tract. Once a blastopore and an archenteron have formed, the embryo is called a gastrula (Gr: gastro = stomach).
Differentiation
At the gastrula stage of development a certain amount of differentiation may be said to have taken place. The cells on the outside of the gastrula constitute the ectoderm (Gr: ecto = outer, derma = skin), and they will develop into the epidermis and the nervous system of the mature animal. The cells of the archenteron constitute the endoderm (Gr: endo = inside), and these will become the lining of the digestive tract and the digestive glands of the fully developed organism. These two primary germ layers will be joined by a third one - the mesoderm.
In sea urchins and sea stars growth, differentiation and morphogenesis continue beyond the gastrula stage until a free-living larval stage, fully capable of moving about and feeding itself, is formed. At a later stage a metamorphosis will occur and the larva will assume the form of the adult starfish or sea urchin.
.
Highly Derived Coelomic and Water-Vascular Morphogenesis in a Starfish with Pelagic Direct Development
D. A. Janies and L. R. McEdward
University of Florida, Department of Zoology, Gainesville, Florida 32611
The coelomic development of the starfish Pteraster tesselatus (order Velatida, family Pterasteridae) is fundamentally different from that reported for all other asteroids. Coeloms arise from seven separate enterocoels that evaginate from different regions of the archenteron. The water-vascular coelomic system develops from the first five enterocoels (homologous to hydrocoel lobes) which extend radially, in a transverse orientation, from the central region of the archenteron. All other coelomic compartments derive from two enterocoels that evaginate later in development from posterior regions of the archenteron. This mode of coelom formation in P. tesselatus leads directly to the adult organization. We hypothesize that this altered pattern of coelomogenesis evolved from the pattern that occurs in the larvae of other spinulosacean asteroids, by a rotation in the site of origin of the anterior enterocoels relative to the archenteron. The altered pattern of coelomogenesis accounts for most of the unusual features of development in P. tesselatus: parallel embryonic and adult axes of symmetry, transverse orientation of the juvenile disk, absence of bilateral symmetry, absence of purely larval structures, and the lack of a metamorphosis. We conclude, contrary to previous interpretations, that P. tesselatus does not have a larval stage and thus represents the only described case of truly direct development in the asteroids.
Ultrastructural study of the hyaline layer of the starfish embryo, Pisaster ochraceus.
Campbell SS, Crawford BJ.
Department of Anatomy, University of British Columbia, Vancouver, Canada.
The hyaline layer (HL) is a multilayered extracellular matrix (ECM) that coats the external surfaces of sea urchin and starfish embryos. It is thought to protect and lubricate the embryo, stabilize the blastomeres during morphogenesis, and regulate nutrient intake. Ultrastructural studies of chemically fixed embryos have shown the HL to consist of two to four sublayers. However, since chemical fixatives may cause collapse and alter the positions and antigenicity of the extracellular components, fixation methods that exclude chemicals may reveal a picture of the HL closer to what is present in vivo. Freeze substitution, a fixation method whereby tissues are rapidly frozen and dehydrated at low temperatures, has proved useful for fixing material rich in ECM. In this study, embryos of the starfish Pisaster ochraceus were fixed for microscopy using freeze substitution and three chemical methods in order to determine, as accurately as possible, the structure of the HL. Embryos appear to be best preserved by freeze substitution and demonstrate a HL consisting of at least six distinct sublayers. Based on staining with anionic dyes, most sublayers appear to contain glycosaminoglycans. Freeze substituted embryos, which were also stained with monoclonal antibodies raised against their ECM, revealed that some molecules are common to all six sublayers, whereas other molecules may be restricted to specific sublayers. This suggests that each sublayer could have a different function. Additional evidence suggests that microvillus associated bodies, present in other marine invertebrate embryos, may anchor the asteroid HL to the cell surface microvilli.
شناسایی مراحل نمو در ستاره دریایی
تسهیم
اولین مرحله ساده که تعدادی از سلولها حاوی اند. مراحل 2 سلولی, 4سلولی, 8 سلولی و الی آخر. این مراحل از اولین , دومین, سومین وتا تسهیمات پایانی حاصل می شوند. سلولهای حاصل از هر مرحله به نام بلاستومر خوانده می شوند. سلولها به سادگی بر تعدادشان افزوده شده تا مراحل 32 سلولی یا 64 سلولی , در این مراحل به جنین مورولا اطلاق می شود. مورولا توده کروی شکل حاصل از 12 تا چند صد سلول است.
بلاستولاسیون
مورولا توده توپر است ولی تسهیم ادامه یافته تا بلاستومر به سمت پیرامون خود کشیده شوند و حفره داخلی پر شده از مایع ایجاد شود. جنین یک توده از سلول هایی است که طویل نیستند ولی تشکیل یک کره را داده اند. در این مرحله جنین بلاستولا خوانده می شود و حفره پر از مایع بلاستوسل خوانده می شود. این تغییرات در حدود 10ساعت پس از لقاح به طول می انجامد. زمانی که بلاستولا برای نخست شکل می گیرد حاوی تقریبا 1000 سلول است.
گاسترولاسیون (لایه زایی)
چند ساعت کوتاه پس از بلاستولا فرایند های گاسترولاسیون (لایه زایی) آغاز می شود. در یک نقطه قطعی از سطح بلاستولا, سلولها شروع به حرکت به سمت مرکز بلاستوسل می کند. در آغاز این شکل گیری منجر به خمیدگی لوله می شود. اگر شما بادکنکی را به آرامی از آب یا هوا پر کنید و با انگشتانتان به داخل بادکنک فشار بیاوریدشما بطور تقریبی گاسرولاسیون را انجام داده اید. سطح باز شدن جنین منفذ بلاستوپور خوانده می شود و در دوتروستومیان ها (دو دهانه ای ها) نظیر توتیا و ستاره دریایی منفذ بلاستوپور یه مخرج آینده موجود تبدیل می شود. لوله خمیده آرکانترون نامیده می شود ودر نهایت به تمام جنین رسیده و در جهت دیگری از سطح جنین باز می شود و اساس شکل گیری لوله گوارشی جانور خواهد شد. از زمانی که منفذ بلاستوپور و ارکانترون شکل گیرد, جنین گاسترولا خوانده می شود.
گاسترولاسیون در جنین میکرولسیتال
در تصاویر ارایه شده، جنین در مرحله گاسترولای اولیه تا مرحله لارو" بی پی ناریا"دیده می شود.روند گاسترولاسیون مانند اندازه و توزیع زرده و تسهیم در بین گونه های مختلف در جزییات متفاوت است. در این مرحله برخی از سلول های سطح جنین به داخل حرکت کرده و لایه های سلولی جدید شکل می گیرد. این حرکات با تمایز سلول های مهاجر بداخل لایه های منفرد بافتی توام است.(ژن های خاصی فعالیت های تمایزی را باعث می شوند.)
در طی لایه زایی یک بلاستولا با خفره بلاستوسل نسبتا بزرگ شکل می گیردکه فضای کافی برای حرکت برخی از سلول های سطحی بداخل جهت تشکیل لایه های جدید را دارد.مشاهدات اولیه ستاره دریای جنین در قطب گیاهی مسطح می شود .(مهاجرت اولیه و تمایز سلول ها) که بداخل بلاستوسل قرورفتگی می یابند. و لایه سلولی داخلی شکل گرفته (هایپوبلاست یا مزودرم )و لایه های مزودرم و اندودرم شکل می گیرند.سلول های سطح خارجی گاسترولا ، اپی بلاست یا اکتودرم می شوند. و حفره بلاستوسل ، گاستروسل یاارکانترون (لوله گوارشی اولیه ) نامیده می شود. که در قطب گیاهی بعنوان بلاستوپور باز می شود.
انتهای داخلی هایپوبلاست (مزودرم) کیسه های مزودرمال را شکل می دهد. این کیسه ها Entrocelic نامیده می شوند. و اعمال انها فعالیت سلومی خوانده می شود. به موازات رشد جنین طی گاسترولاسیون ، سلول هایی از هایپوبلاست جوانه زده و برای تشکیل سلول های مزانشیمی به داخل مهاجرت میکنند.
تمایز
در مرحله گاسترولا , اندازه قطعی از تمایز نمو می یابد. سلول های سطح بیرونی گاسترولا اکتو درم را تشکیل می دهد و در آنها اپیدرم و سیستم عصبی جانور بالغ تمایز می یابد. این دولایه بوسیله لایه سوم ، مزودرم به همدیگر متصل می شوند.
در توتیا و ستاره دریایی رشد ،تمایز و شکل گیری در گاسترولا ادامه می یابد. مرحله لارو آزاد که بطور کامل قادر به حرکت و تغذیه از محیط اطراف خود است، شکل میگیرد. در مرحله بعد یک دگردیسی رخ داده و لارو به ستاره دریایی بالغ یا توتیای بالغ تبدیل می شود
تصاویری از تسهیم و لایه زایی ستاره دریایی
این صفحه یکسری از تصاویری که از تخم میکرو لیستال از سلول لقاح نیافته با مرحله بلاستولادر جنین ستاره دریایی را نشان می دهد. تخم های میکرو لیستال دارای اندازه خیلی کوچکی از زرده ذخیره شده اند. در نتیجه وجود اندازه کوچک زرده ,این حیوانات یکی از دو روش نمو را نشان میدهد.
در اولین روش لارو در مرحله جنینی از مواد تغذیه ای محیط تغذیه می شود. (در جنین های آورده شده زیر) در روش دوم که نمو داخلی است. نیاز های اساسی به وسیله مادر برآورده می شود(چیزی که ما در پستانداران مشاهده می کنیم.)
Later
تخم لقاح نیافته که از طریق اندازه و هسته مشخص (پیکان بزرگ) با هستک مشخص (پیکان کوچک) و فقدان غشای لقاحی قابل تشخیص است. تصویر زیگوت یا تخم لقاح که بوسیله غشای لقاحی (پیکانها) احاطه کرده سلول و فضای دور ویتیلینی که با مایع پر شده است،مشخص است.
Later
مرحله دو سلولی که زیگوت اولین تسهیم را کامل کرده است و تسهیم اول کامل و مساوی است.(نظیر تخمک کامل که به دو سلول تقسیم می شود). تسهیم از حول نباتی-جانوری عبور کرده است و دو بلاستومر مشابه تولید می کند. مرحله چهار سلولی که تسهیم دوم معمولا از سراسر محور گیاهی-نباتی عبور می کند. ولی بر تسهیم اول عمود است. در پایان چهار بلاستومر کامل با اندازه مساوی ایجاد می شود.
Later
مرحله هشت سلولی, سومین تسهیم در سطح استوایی رخ میدهد.(عمود بر دو تسهیم اول و محور نباتی-جانوری). چهار بلاستومر فوقانی (قطب جانوری) در اندازه کوچکتر از چهار بلاستومر تحتانی (قطب گیاهی) هستند. مرحله 32 سلولی: پس از مرحله 16 سلولی تقسیم بطور پیچیده دنبال می گردد. افزایش تعداد سلولها و تقسیمات بلاستومرها همزمان انجام می شود. تقسیم ادامه یافته و یک توده سلولی که خودش را به بلاستولا سازماندهی می کند, شکل می گیرد.
Later
بلاستولای اولیه: تسهیم ادامه پیدا می کند. در مرکز, تماس سلول ها با یکدیگر از بین رفته, و حفره مرکز پوشیده از مایع (بلاستوسل) شکل می گیرد. این بلاستوسل توسط یک لایه از سلول احاطه می شود. کره توخالی شکل گرفته به عنوان بلاستولا شناخته می شود. بلاستولای تاخیری: مشابه بلاستولای اولیه توسط یک لایه منفرد از سلول های احاطه کننده حفره مرکزی قابل تشخیص است. بلاستومرها کوچک و بطور مشخص واضح نیستند.بلاستومرهای قطب گیاهی vp ضخیم تر از قطب جانوری APظاهر می شود.
Later
گاسترولای اولیه
animal pole (AP)
vegetal pole (VP)
Later
گاسترولای میانی گاسترولای تاخیری
C
O
L
O
U
R
I
Z
E
!!
لارو بی پی ناریا
1. Blastocoel
2. Enterocoelic pouch
3. Mouth
4. Archenteron
5. Blastopore (anus) • blue=Ectoderm
• red=Mesoderm
• yellow=Endoderm
.
This page shows a series of photographs which follows a microlecithal (oligolecithal) egg from the unfertilized single cell through to the late blastula. Microlecithal eggs have a very small amount of stored yolk, distributed evenly throughout the cytoplasm. As a result of this small amount of yolk, these animals show one of two different methods of development. The first is to have a feeding larval stage (as this embryo will have) to meet the nutritional requirements of the embryo, with the second being to have internal development such that the nutritional requirements of the embryo are met by the mother (what we will see later with mammals). These particular slides are whole mounts of starfish embryos.
Later
This picture is of the unfertilized egg. It can be differentiated from the zygote by the presence of a large, conspicuous nucleus (large arrow) with obvious nucleolus (smaller arrow) and by the lack of a fertilization membrane. This shows the zygote (fertilized cell). It is recognized by the presence of the fertilization membrane (arrows) surrounding it and the peripheral, fluid-filled perivitelline space.
Later
This is the two cell stage. By this stage, the zygote has completed its first cleavage, which is both equal and holoblastic (i.e. the entire ovum is divided into cells). The division (cleavage) has passed through the animal-vegetal axis, producing two similar blastomeres. This is the four cell stage. This second cleavage also passes through the animal-vegetal axis, but perpendicular to the first cleavage (described previously). Four equal-sized blastomeres are the end result.
Later
The eight cell stage. Here, the third cleavage has occurred in the equatorial plane (perpendicular to the first two cleavages and the animal-vegetal axis). Note that the upper four blastomeres (the animal pole) are slightly smaller than the lower four blastomeres (the vegetal pole). NOTE: The sixteen cell stage is not shown - its division is once again vertical, along the animal-vegetal axis. The thirty-two cell stage is seen here. After the 16 cell stage, the cleavages become more difficult to follow, due to the increasing number of cells and to the division of blastomeres becoming asynchronous. Cleavage continues, forming a mass of cells which organizes itself into the blastula. The lighter area in the centre of the embryo is the beginning of the blastocoel.
Later
This shows the early blastula. With continuing cleavage, the cells in centre begin to lose contact with one another, and a central fluid-filled cavity (the blastocoel) forms. This blastocoel is surrounded by a single layer of cells, forming the hollow sphere know as the blastula. This shows the late blastula. Like the early blastula, it is characterized by a single layer of cells surrounding the central hollow area - the blastocoel (B). The blastomeres are seen to be smaller and are individually not as obvious. The blastomeres at the vegetal pole (VP) are taller than those at the animal pole (AP), making the vegetal pole appear slightly thicker.
Gastrulation in the Microlecithal Embryo
On this page, a series of photographs can be seen which follow a microlecithal (oligolecithal) embryo from the early gastrula stage through gastrulation to the early bipinnaria larval stage.
The process of gastrulation is affected, as was cleavage, by the amount and the distribution of yolk. As a result, gastrulation differs between different species in details, but the principle is the same in all these species. During gastrulation, some of the cells from the surface of the embryo move to the interior and replicate, thereby forming new layers of cells. These movements are coupled with the differentiation of the migrating cells (caused by the differential activity of certain genes) into histologically unique layers.
During microlecithal gastrulation, a blastula with a relatively large blastocoel is usually formed. As a result, there is plenty of space to move some of the surface cells to the inside to form new layers. This is first seen as the starfish embryo becomes flattened at the vegetal pole (the initial migration and differentiation of cells), which will then invaginate within the blastocoel. When this occurs, the internal layer of cells formed (called the hypoblast or mesendoderm) are the cells which gives rise to the endoderm and mesoderm germ layers. The cells who remain on the outer surface of the gastrula become the epiblast (ectoderm). The cavity created by the invagination is called the gastrocoel or archenteron (primitive gut), whose opening at the vegetal pole is called the blastopore. The anterior end of the hypoblast (mesendoderm) develops out-pocketings (evaginations) on each side to form the mesoderm (mesodermal pouches). These pouches are also called enterocoelic pouches and the cavities within them are called the coelomic cavities. As the embryo grows during gastrulation, cells also bud off the hypoblast and migrate into the blastocoel to form mesenchymal cells.
Later
This is the early starfish gastrula. The vegetal pole (VP) is showing the first signs of invagination. AP=animal pole, VP=vegetal pole. This shows the early gastrula. The beginning of the invagination which will form the gastrocoel (a.k.a. archenteron) is obvious now, as it invaginates anteriorly (toward the animal pole (AP)) from the vegetal pole (VP), where the blastopore is formed.
Later
Here we see the mid gastrula. The gastrocoel continues to develop as the embryo grows. AP=animal pole, VP=vegetal pole. This is the late gastrula. This picture is a dorsal view of the embryo, with anterior being toward the animal pole (AP) and posterior toward the vegetal pole (VP). Note the beginning of the formation of enterocoelic pouches on the right and left side of the anterior end of the gastrocoel.
C
O
L
O
U
R
I
Z
E
!!
This is the early bipinnaria larva seen in lateral view (from the side). The dorsal aspect of the embryo is facing right and the ventral aspect is on the left side of this picture. The blastopore is at the posterior end (bottom) and is now developing into the anus, while the anterior end of the gastrocoel has bent toward the ventral aspect of the embryo, where a mouth has formed. Here we see the same picture as on the right, but now labelled and in glorious technicolour!
Blastocoel
Enterocoelic pouch
Mouth
Archenteron
Blastopore (anus) blue=Ectoderm
red=Mesoderm
yellow=Endoderm
Cleavage and Morphogenesis
Development of Sea Star Embryos (Rust, fig. 110)
Egg and Zygote
The unfertilized egg (1a) has a large and conspicuous nucleus. Compare the amount of yolk and cytoplasm to the size of the nucleus. The nucleus of the zygote (1b) is less conspicuous. The fertilization membrane is not present in this preserved material
.
cell stage
Successive cell divisions divide the zygote into smaller and more numerous cells. Compare the size of the 2,4 and 8 cell stages with the size of the original zygote. Note the cleavage furrows formed by the pinching in of the cell membranes during division. From the arrangement of the cells determine the plane of cleavage that produced the cells.
.
Early blastula
Cleavage continues producing many additional cells which arrange themselves around a fluid filled cavity (blastocoel). The blastula (7) begins to take on the appearance of a ball.
. Blastula
This ball-like stage is called the blastula (5). Compare the size of the blastula to the original zygote. Note the blastocoel (6).
late gastrula
During the late blastula stage a few cells on one side of the developing embryo begin to push into the blastocoel (8). When this process (gastrulation) becomes visible as an indentation, the developing embryo is termed a gastrula (9). The point of indentation is termed the blastopore (10). As the invagination process continues (11),with more cells pushing into the interior, a new cell layer is formed and a new cavity (12) (archenteron) appears within this new layer of cells. This cavity opens through the blastopore (14) to the outside of the embryo. Later a third layer of cells develops between the original two layers. These three cell layers (ectoderm, endoderm and mesoderm) provide the "bricks and mortar" for the differentiation of the embryo's organ systems.
. Larval
The developmental process to the end of the gastrula (13) stage takes approximately two days. At this time the gastrula transforms itself into a free swimming (via cilia bands) (18) and feeding larva (15). This change includes the formation of a digestive system with two openings (mouth (17) and anus (16)). The larva grows as it feeds on algae and diatoms. After several months the larva undergoes a process called metamorphosis that transforms it into a tiny sea star.
Sea stars belong to the Phylum Echinodermata, and display the typical deuterostome pattern of early development. In sea stars, fertilization and development take place externally and the eggs carry little yolk, so it is easy to observe early development. Obtain a slide of developing sea star embryos and
identify the following developmental stages.
_Cleavage
The earliest stages are simply named for the number of cells they contain: the 2-cell stage, the 4-cell stage, the 8-cell stage, and so on. These stages result from the first cleavage, the second cleavage, the third cleavage, etc. (Individual cells that form during these cleavages are called blastomeres (Gr: blastus = a bud, mere = part).) Once the cells become too numerous to be counted easily, which usually happens around the 32- or 64-cell stage, this terminology is abandoned and the embryo is said to be in the morula (Latin: little mulberry) stage. The morula is a spherical mass of from a few dozen to several hundred cells.
Blastulation
The morula is solid, but as cleavage continues the blastomeres move toward the periphery of the mass, leaving a fluid-filled cavity in the interior. Soon the embryo no longer resembles a simple mass of cells, but instead takes on the form of a sphere. In this stage the embryo is called a blastula (little bud), and the fluid-filled cavity is called a blastocoel (Gr: coelo = hollow). This change takes place about ten hours after fertilization. When the blastula first forms it contains about 1000 cells.
Gastrulation
A few hours after the formation of the blastula the process of gastrulation begins. At a certain point on the surface of the blastula cells begin to move toward the center of the blastocoel, so that an opening is formed leading into a blind or dead-end tube. If you were to take a rubber balloon loosely filled with water or air and poke your finger into it without breaking the balloon you would be at least roughly approximating the process of gastrulation. The opening on the surface of the embryo is called the blastopore, and in deuterostomes such as sea urchins and sea stars this will become the anus of the mature organism. The blind or dead-end tube is called the archenteron (Gr: archi = first, enterum = intestine). This will eventually reach all the way through and open out the other side, and thus will form the basis of the animal's digestive tract. Once a blastopore and an archenteron have formed, the embryo is called a gastrula (Gr: gastro = stomach).
Differentiation
At the gastrula stage of development a certain amount of differentiation may be said to have taken place. The cells on the outside of the gastrula constitute the ectoderm (Gr: ecto = outer, derma = skin), and they will develop into the epidermis and the nervous system of the mature animal. The cells of the archenteron constitute the endoderm (Gr: endo = inside), and these will become the lining of the digestive tract and the digestive glands of the fully developed organism. These two primary germ layers will be joined by a third one - the mesoderm.
In sea urchins and sea stars growth, differentiation and morphogenesis continue beyond the gastrula stage until a free-living larval stage, fully capable of moving about and feeding itself, is formed. At a later stage a metamorphosis will occur and the larva will assume the form of the adult starfish or sea urchin.
.
Highly Derived Coelomic and Water-Vascular Morphogenesis in a Starfish with Pelagic Direct Development
D. A. Janies and L. R. McEdward
University of Florida, Department of Zoology, Gainesville, Florida 32611
The coelomic development of the starfish Pteraster tesselatus (order Velatida, family Pterasteridae) is fundamentally different from that reported for all other asteroids. Coeloms arise from seven separate enterocoels that evaginate from different regions of the archenteron. The water-vascular coelomic system develops from the first five enterocoels (homologous to hydrocoel lobes) which extend radially, in a transverse orientation, from the central region of the archenteron. All other coelomic compartments derive from two enterocoels that evaginate later in development from posterior regions of the archenteron. This mode of coelom formation in P. tesselatus leads directly to the adult organization. We hypothesize that this altered pattern of coelomogenesis evolved from the pattern that occurs in the larvae of other spinulosacean asteroids, by a rotation in the site of origin of the anterior enterocoels relative to the archenteron. The altered pattern of coelomogenesis accounts for most of the unusual features of development in P. tesselatus: parallel embryonic and adult axes of symmetry, transverse orientation of the juvenile disk, absence of bilateral symmetry, absence of purely larval structures, and the lack of a metamorphosis. We conclude, contrary to previous interpretations, that P. tesselatus does not have a larval stage and thus represents the only described case of truly direct development in the asteroids.
Ultrastructural study of the hyaline layer of the starfish embryo, Pisaster ochraceus.
Campbell SS, Crawford BJ.
Department of Anatomy, University of British Columbia, Vancouver, Canada.
The hyaline layer (HL) is a multilayered extracellular matrix (ECM) that coats the external surfaces of sea urchin and starfish embryos. It is thought to protect and lubricate the embryo, stabilize the blastomeres during morphogenesis, and regulate nutrient intake. Ultrastructural studies of chemically fixed embryos have shown the HL to consist of two to four sublayers. However, since chemical fixatives may cause collapse and alter the positions and antigenicity of the extracellular components, fixation methods that exclude chemicals may reveal a picture of the HL closer to what is present in vivo. Freeze substitution, a fixation method whereby tissues are rapidly frozen and dehydrated at low temperatures, has proved useful for fixing material rich in ECM. In this study, embryos of the starfish Pisaster ochraceus were fixed for microscopy using freeze substitution and three chemical methods in order to determine, as accurately as possible, the structure of the HL. Embryos appear to be best preserved by freeze substitution and demonstrate a HL consisting of at least six distinct sublayers. Based on staining with anionic dyes, most sublayers appear to contain glycosaminoglycans. Freeze substituted embryos, which were also stained with monoclonal antibodies raised against their ECM, revealed that some molecules are common to all six sublayers, whereas other molecules may be restricted to specific sublayers. This suggests that each sublayer could have a different function. Additional evidence suggests that microvillus associated bodies, present in other marine invertebrate embryos, may anchor the asteroid HL to the cell surface microvilli.
شناسایی مراحل نمو در ستاره دریایی
تسهیم
اولین مرحله ساده که تعدادی از سلولها حاوی اند. مراحل 2 سلولی, 4سلولی, 8 سلولی و الی آخر. این مراحل از اولین , دومین, سومین وتا تسهیمات پایانی حاصل می شوند. سلولهای حاصل از هر مرحله به نام بلاستومر خوانده می شوند. سلولها به سادگی بر تعدادشان افزوده شده تا مراحل 32 سلولی یا 64 سلولی , در این مراحل به جنین مورولا اطلاق می شود. مورولا توده کروی شکل حاصل از 12 تا چند صد سلول است.
بلاستولاسیون
مورولا توده توپر است ولی تسهیم ادامه یافته تا بلاستومر به سمت پیرامون خود کشیده شوند و حفره داخلی پر شده از مایع ایجاد شود. جنین یک توده از سلول هایی است که طویل نیستند ولی تشکیل یک کره را داده اند. در این مرحله جنین بلاستولا خوانده می شود و حفره پر از مایع بلاستوسل خوانده می شود. این تغییرات در حدود 10ساعت پس از لقاح به طول می انجامد. زمانی که بلاستولا برای نخست شکل می گیرد حاوی تقریبا 1000 سلول است.
گاسترولاسیون (لایه زایی)
چند ساعت کوتاه پس از بلاستولا فرایند های گاسترولاسیون (لایه زایی) آغاز می شود. در یک نقطه قطعی از سطح بلاستولا, سلولها شروع به حرکت به سمت مرکز بلاستوسل می کند. در آغاز این شکل گیری منجر به خمیدگی لوله می شود. اگر شما بادکنکی را به آرامی از آب یا هوا پر کنید و با انگشتانتان به داخل بادکنک فشار بیاوریدشما بطور تقریبی گاسرولاسیون را انجام داده اید. سطح باز شدن جنین منفذ بلاستوپور خوانده می شود و در دوتروستومیان ها (دو دهانه ای ها) نظیر توتیا و ستاره دریایی منفذ بلاستوپور یه مخرج آینده موجود تبدیل می شود. لوله خمیده آرکانترون نامیده می شود ودر نهایت به تمام جنین رسیده و در جهت دیگری از سطح جنین باز می شود و اساس شکل گیری لوله گوارشی جانور خواهد شد. از زمانی که منفذ بلاستوپور و ارکانترون شکل گیرد, جنین گاسترولا خوانده می شود.
گاسترولاسیون در جنین میکرولسیتال
در تصاویر ارایه شده، جنین در مرحله گاسترولای اولیه تا مرحله لارو" بی پی ناریا"دیده می شود.روند گاسترولاسیون مانند اندازه و توزیع زرده و تسهیم در بین گونه های مختلف در جزییات متفاوت است. در این مرحله برخی از سلول های سطح جنین به داخل حرکت کرده و لایه های سلولی جدید شکل می گیرد. این حرکات با تمایز سلول های مهاجر بداخل لایه های منفرد بافتی توام است.(ژن های خاصی فعالیت های تمایزی را باعث می شوند.)
در طی لایه زایی یک بلاستولا با خفره بلاستوسل نسبتا بزرگ شکل می گیردکه فضای کافی برای حرکت برخی از سلول های سطحی بداخل جهت تشکیل لایه های جدید را دارد.مشاهدات اولیه ستاره دریای جنین در قطب گیاهی مسطح می شود .(مهاجرت اولیه و تمایز سلول ها) که بداخل بلاستوسل قرورفتگی می یابند. و لایه سلولی داخلی شکل گرفته (هایپوبلاست یا مزودرم )و لایه های مزودرم و اندودرم شکل می گیرند.سلول های سطح خارجی گاسترولا ، اپی بلاست یا اکتودرم می شوند. و حفره بلاستوسل ، گاستروسل یاارکانترون (لوله گوارشی اولیه ) نامیده می شود. که در قطب گیاهی بعنوان بلاستوپور باز می شود.
انتهای داخلی هایپوبلاست (مزودرم) کیسه های مزودرمال را شکل می دهد. این کیسه ها Entrocelic نامیده می شوند. و اعمال انها فعالیت سلومی خوانده می شود. به موازات رشد جنین طی گاسترولاسیون ، سلول هایی از هایپوبلاست جوانه زده و برای تشکیل سلول های مزانشیمی به داخل مهاجرت میکنند.
تمایز
در مرحله گاسترولا , اندازه قطعی از تمایز نمو می یابد. سلول های سطح بیرونی گاسترولا اکتو درم را تشکیل می دهد و در آنها اپیدرم و سیستم عصبی جانور بالغ تمایز می یابد. این دولایه بوسیله لایه سوم ، مزودرم به همدیگر متصل می شوند.
در توتیا و ستاره دریایی رشد ،تمایز و شکل گیری در گاسترولا ادامه می یابد. مرحله لارو آزاد که بطور کامل قادر به حرکت و تغذیه از محیط اطراف خود است، شکل میگیرد. در مرحله بعد یک دگردیسی رخ داده و لارو به ستاره دریایی بالغ یا توتیای بالغ تبدیل می شود
تصاویری از تسهیم و لایه زایی ستاره دریایی
این صفحه یکسری از تصاویری که از تخم میکرو لیستال از سلول لقاح نیافته با مرحله بلاستولادر جنین ستاره دریایی را نشان می دهد. تخم های میکرو لیستال دارای اندازه خیلی کوچکی از زرده ذخیره شده اند. در نتیجه وجود اندازه کوچک زرده ,این حیوانات یکی از دو روش نمو را نشان میدهد.
در اولین روش لارو در مرحله جنینی از مواد تغذیه ای محیط تغذیه می شود. (در جنین های آورده شده زیر) در روش دوم که نمو داخلی است. نیاز های اساسی به وسیله مادر برآورده می شود(چیزی که ما در پستانداران مشاهده می کنیم.)
Later
تخم لقاح نیافته که از طریق اندازه و هسته مشخص (پیکان بزرگ) با هستک مشخص (پیکان کوچک) و فقدان غشای لقاحی قابل تشخیص است. تصویر زیگوت یا تخم لقاح که بوسیله غشای لقاحی (پیکانها) احاطه کرده سلول و فضای دور ویتیلینی که با مایع پر شده است،مشخص است.
Later
مرحله دو سلولی که زیگوت اولین تسهیم را کامل کرده است و تسهیم اول کامل و مساوی است.(نظیر تخمک کامل که به دو سلول تقسیم می شود). تسهیم از حول نباتی-جانوری عبور کرده است و دو بلاستومر مشابه تولید می کند. مرحله چهار سلولی که تسهیم دوم معمولا از سراسر محور گیاهی-نباتی عبور می کند. ولی بر تسهیم اول عمود است. در پایان چهار بلاستومر کامل با اندازه مساوی ایجاد می شود.
Later
مرحله هشت سلولی, سومین تسهیم در سطح استوایی رخ میدهد.(عمود بر دو تسهیم اول و محور نباتی-جانوری). چهار بلاستومر فوقانی (قطب جانوری) در اندازه کوچکتر از چهار بلاستومر تحتانی (قطب گیاهی) هستند. مرحله 32 سلولی: پس از مرحله 16 سلولی تقسیم بطور پیچیده دنبال می گردد. افزایش تعداد سلولها و تقسیمات بلاستومرها همزمان انجام می شود. تقسیم ادامه یافته و یک توده سلولی که خودش را به بلاستولا سازماندهی می کند, شکل می گیرد.
Later
بلاستولای اولیه: تسهیم ادامه پیدا می کند. در مرکز, تماس سلول ها با یکدیگر از بین رفته, و حفره مرکز پوشیده از مایع (بلاستوسل) شکل می گیرد. این بلاستوسل توسط یک لایه از سلول احاطه می شود. کره توخالی شکل گرفته به عنوان بلاستولا شناخته می شود. بلاستولای تاخیری: مشابه بلاستولای اولیه توسط یک لایه منفرد از سلول های احاطه کننده حفره مرکزی قابل تشخیص است. بلاستومرها کوچک و بطور مشخص واضح نیستند.بلاستومرهای قطب گیاهی vp ضخیم تر از قطب جانوری APظاهر می شود.
Later
گاسترولای اولیه
animal pole (AP)
vegetal pole (VP)
Later
گاسترولای میانی گاسترولای تاخیری
C
O
L
O
U
R
I
Z
E
!!
لارو بی پی ناریا
1. Blastocoel
2. Enterocoelic pouch
3. Mouth
4. Archenteron
5. Blastopore (anus) • blue=Ectoderm
• red=Mesoderm
• yellow=Endoderm
.
هیچ نظری موجود نیست:
ارسال یک نظر