


[edit] Sea urchins
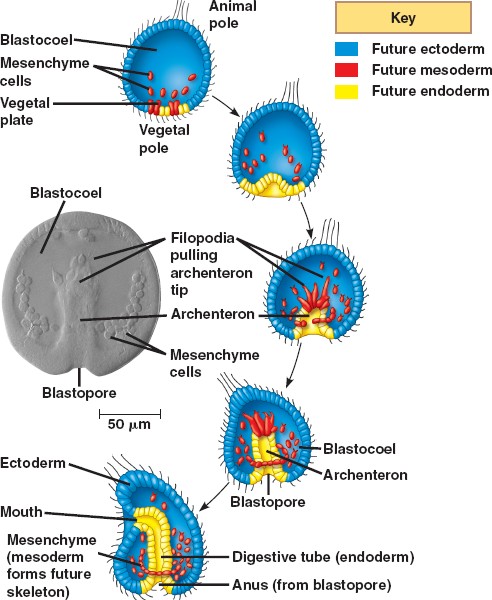
The following description concerns gastrulation in echinoderms, representative of the triploblasts, or animals with three embryonic germ layers. The illustration, however, depicts the gastrulation of a diploblast, animals with two germ layers.
Sea urchins deviate from simple cleavage at the fourth cleavage. The four vegetal blastomeres divide unequally to produce four micromeres at the vegetal pole and four macromeres in the middle of the embryo. The animal cells divide meridionally and produce mesomeres.
At the beginning of vertebrate gastrulation, the embryo is a hollow ball of cells known as the blastula, with an animal pole and a vegetal pole. The vegetal pole begins to flatten to form the vegetal plate. Some of the cells of the vegetal pole detach and through ingression become primary mesenchyme cells. The mesenchyme cells divide rapidly and migrate along the extracellular matrix (basal lamina) to different parts of the blastocoel. The migration is believed to be dependent upon sulfated proteoglycans on the surface of the cells and molecules on the basal lamina such as fibronectin. The cells move by forming filopodia that identify the specific target location. These filopodia then organize into syncytial cables that deposit the calcium carbonate that makes up the spicules (the skeleton of the pluteus larva).
During the second phase of gastrulation, the vegetal plate invaginates into the interior, replacing the blastocoelic cavity and thereby forming a new cavity, the archenteron (literally: primitive gut), the opening into which is the blastopore. The arechenteron is elongated by three mechanisms.
First, the initial invagination is caused by a differential expansion of the inner layer made of fibropellins and outer layer made of hyalin to cause the layers to bend inward.
Second, the archenteron is formed through convergent extension. Convergent extension results when cells intercalate to narrow the tissue and move it forward.
Third, secondary mesenchyme pull the tip of the archenteron towards the animal pole. Secondary mesenchyme are formed from cells that ingress from, but remain attached to, the roof of the archenteron. These cells extend filopodia that use guidance cues to find the future mouth region. Upon reaching the target site, the cells contract to pull the archenteron to fuse with the ectoderm. Once the archenteron reaches the animal pole, a perforation forms, and the archenteron becomes a digestive tract passing all the way through the embryo.
The three embryonic germ layers have now formed. The endoderm, consisting of the archenteron, will develop into the digestive tract. The ectoderm, consisting of the cells on the outside of the gastrula that played little part in gastrulation, will develop into the skin and the central nervous system. The mesoderm, consisting of the mesenchyme cells that have proliferated in the blastocoel, will become all the other internal organs.
An extracellular matrix molecule that is selectively expressed during development is important for gastrulation in the sea urchin embryo.
Berg LK, Chen SW, Wessel GM.
Department of Molecular and Cell Biology & Biochemistry, Brown University, Providence, RI 02912, USA.
The extracellular matrix is important in the regulation of many cellular events of early development including migration, shape change, proliferation and gene expression. In the sea urchin embryo, disruption of the extracellular matrix results in selective defects in each of these events during gastrulation. Here we describe a new molecule of the extracellular matrix in Lytechinus variegatus, referred to as ECM 18, that has several important features. First, antibody interference of ECM 18 results in a profound but reversible inhibition of primary mesenchyme cell organization and endoderm morphogenesis during gastrulation. Second, during gastrulation, ECM 18 mRNA accumulates to highest levels in the invaginating endoderm and the ECM 18 protein deposited in the basal lamina surrounding the archenteron as well as in other areas of the blastocoel wall. Immunolocalization by fluorescence and electron microscopy demonstrates the selective accumulation of ECM 18 in the extracellular matrix. Third, although the mRNA encoding ECM 18 is present throughout development, the protein accumulates only during gastrulation. ECM 18 protein is not detected in eggs or early embryos and analysis of polysome-associated mRNA suggests that at least part of the translational regulation of ECM 18 is at the level of ECM 18 mRNA-polysome formation. Finally, sequence analysis of ECM 18 shows that the protein contains a repeat sequence with a conserved cysteine motif, suggestive of involvement in protein-protein interactions. Thus, ECM 18 appears to be important in mediating select morphogenetic changes during gastrulation and the pattern of its expression in the embryo is unique among the extracellular matrix molecules known in this embryo.
PMID: 8625821 [PubMed - indexed for MEDLINE]
transcription Factor Activation in the Sea Urchin Embryo
It should be noted that the biochemical nature of the vegetalizing signal(s) has not yet been elucidated. Hörstadius interpreted these experiments in terms of gradients of animalizing and vegetalizing substances. More recently, Davidson (1989) sought to explain these phenomena in terms of transcriptional regulatory proteins (such as those that bind to promoters or enhancers) that are localized throughout the egg and that become activated by inductive interactions between adjacent cells. One of these factors, the one in the micromere precursors of the primary mesenchyme cells, is thought to become active autonomously, early in cleavage. This factor would cause the determination of the micromeres, alter their cell membranes so they could react with the cells above them, and initiate the cascade of determination. By the end of blastulation, there would be five territories that would constitute both the precursor cells for certain regions of the pluteus larva and the regions of specific gene activation. For example, the cytoskeletal actin gene, which is expressed only in the aboral ectoderm cells, cannot be activated anywhere else in the embryo. Similarly, if one injects the gene for the skeletal matrix protein anywhere else in the embryo except the region of skeletogenic mesenchyme, it will not be activated (Hough-Evans et al., 1987; Sucov et al., 1988). Therefore, each territory would constitute a region of differential gene expression that would be reflected in different parts of the organism being formed.
However, if the embryo is perturbed, the neighbors would differ. New fates would ensue as the new vegetal cells activated their skeletogenic mesenchyme determinants and started activation of their neighbors. To see if this was indeed the case, Ransick and Davidson (1993) transplanted micromeres into the animal cap of an 8-cell sea urchin embryo. These skeletogenic mesenchyme precursor cells induced the formation of the archenteron and the expression of the archenteron-specific genes. The fluorescently labeled micromeres did not contribute structurally to the secondary archenteron, but provided a short-range inductive signal that respecified the animal cap cells.
Literature Cited
Davidson, E. H. 1989. Lineage-specific gene expression and the regulative capacities of the sea urchin embryo: a proposed mechanism. Development 105: 421-446.
Hough-Evans, B. R., Franks, R. R., Cameron, R. A., Britten, R. J. and Davidson, E. H. 1987. Correct cell type-specific expression of a fusion gene injected into sea urchin eggs. Dev. Biol. 121: 576-579.
Ransick, A. and Davidson, E. H. 1993. A complete second gut induced by transplanted micromeres in the sea urchin embryo. Science 259: 1134-1138.
Sucov, H. M., Hough-Evans, B. R., Franks, R. R., Britten, R. J. and Davidson, E. H. 1988. A regulatory domain that directs lineage-specific expression of a skeletal matrix protein in the sea urchin embryo. Genes Dev. 2: 1238-1250.
© All the material on this website is protected by copyright. It may not be reproduced in any form without permission from the copyright holder.
HOME :: CHAPTER 8 :: SEA URCHIN CELL SPECIFICATION :: TRANSCRIPTION FACTOR ACTIVATION IN THE SEA URCHIN EMBRYO PREVIOUS :: NEXT
Home Link
The following description concerns gastrulation in echinoderms, representative of the triploblasts, or animals with three embryonic germ layers. The illustration, however, depicts the gastrulation of a diploblast, animals with two germ layers.
Sea urchins deviate from simple cleavage at the fourth cleavage. The four vegetal blastomeres divide unequally to produce four micromeres at the vegetal pole and four macromeres in the middle of the embryo. The animal cells divide meridionally and produce mesomeres.
At the beginning of vertebrate gastrulation, the embryo is a hollow ball of cells known as the blastula, with an animal pole and a vegetal pole. The vegetal pole begins to flatten to form the vegetal plate. Some of the cells of the vegetal pole detach and through ingression become primary mesenchyme cells. The mesenchyme cells divide rapidly and migrate along the extracellular matrix (basal lamina) to different parts of the blastocoel. The migration is believed to be dependent upon sulfated proteoglycans on the surface of the cells and molecules on the basal lamina such as fibronectin. The cells move by forming filopodia that identify the specific target location. These filopodia then organize into syncytial cables that deposit the calcium carbonate that makes up the spicules (the skeleton of the pluteus larva).
During the second phase of gastrulation, the vegetal plate invaginates into the interior, replacing the blastocoelic cavity and thereby forming a new cavity, the archenteron (literally: primitive gut), the opening into which is the blastopore. The arechenteron is elongated by three mechanisms.
First, the initial invagination is caused by a differential expansion of the inner layer made of fibropellins and outer layer made of hyalin to cause the layers to bend inward.
Second, the archenteron is formed through convergent extension. Convergent extension results when cells intercalate to narrow the tissue and move it forward.
Third, secondary mesenchyme pull the tip of the archenteron towards the animal pole. Secondary mesenchyme are formed from cells that ingress from, but remain attached to, the roof of the archenteron. These cells extend filopodia that use guidance cues to find the future mouth region. Upon reaching the target site, the cells contract to pull the archenteron to fuse with the ectoderm. Once the archenteron reaches the animal pole, a perforation forms, and the archenteron becomes a digestive tract passing all the way through the embryo.
The three embryonic germ layers have now formed. The endoderm, consisting of the archenteron, will develop into the digestive tract. The ectoderm, consisting of the cells on the outside of the gastrula that played little part in gastrulation, will develop into the skin and the central nervous system. The mesoderm, consisting of the mesenchyme cells that have proliferated in the blastocoel, will become all the other internal organs.
An extracellular matrix molecule that is selectively expressed during development is important for gastrulation in the sea urchin embryo.
Berg LK, Chen SW, Wessel GM.
Department of Molecular and Cell Biology & Biochemistry, Brown University, Providence, RI 02912, USA.
The extracellular matrix is important in the regulation of many cellular events of early development including migration, shape change, proliferation and gene expression. In the sea urchin embryo, disruption of the extracellular matrix results in selective defects in each of these events during gastrulation. Here we describe a new molecule of the extracellular matrix in Lytechinus variegatus, referred to as ECM 18, that has several important features. First, antibody interference of ECM 18 results in a profound but reversible inhibition of primary mesenchyme cell organization and endoderm morphogenesis during gastrulation. Second, during gastrulation, ECM 18 mRNA accumulates to highest levels in the invaginating endoderm and the ECM 18 protein deposited in the basal lamina surrounding the archenteron as well as in other areas of the blastocoel wall. Immunolocalization by fluorescence and electron microscopy demonstrates the selective accumulation of ECM 18 in the extracellular matrix. Third, although the mRNA encoding ECM 18 is present throughout development, the protein accumulates only during gastrulation. ECM 18 protein is not detected in eggs or early embryos and analysis of polysome-associated mRNA suggests that at least part of the translational regulation of ECM 18 is at the level of ECM 18 mRNA-polysome formation. Finally, sequence analysis of ECM 18 shows that the protein contains a repeat sequence with a conserved cysteine motif, suggestive of involvement in protein-protein interactions. Thus, ECM 18 appears to be important in mediating select morphogenetic changes during gastrulation and the pattern of its expression in the embryo is unique among the extracellular matrix molecules known in this embryo.
PMID: 8625821 [PubMed - indexed for MEDLINE]
transcription Factor Activation in the Sea Urchin Embryo
It should be noted that the biochemical nature of the vegetalizing signal(s) has not yet been elucidated. Hörstadius interpreted these experiments in terms of gradients of animalizing and vegetalizing substances. More recently, Davidson (1989) sought to explain these phenomena in terms of transcriptional regulatory proteins (such as those that bind to promoters or enhancers) that are localized throughout the egg and that become activated by inductive interactions between adjacent cells. One of these factors, the one in the micromere precursors of the primary mesenchyme cells, is thought to become active autonomously, early in cleavage. This factor would cause the determination of the micromeres, alter their cell membranes so they could react with the cells above them, and initiate the cascade of determination. By the end of blastulation, there would be five territories that would constitute both the precursor cells for certain regions of the pluteus larva and the regions of specific gene activation. For example, the cytoskeletal actin gene, which is expressed only in the aboral ectoderm cells, cannot be activated anywhere else in the embryo. Similarly, if one injects the gene for the skeletal matrix protein anywhere else in the embryo except the region of skeletogenic mesenchyme, it will not be activated (Hough-Evans et al., 1987; Sucov et al., 1988). Therefore, each territory would constitute a region of differential gene expression that would be reflected in different parts of the organism being formed.
However, if the embryo is perturbed, the neighbors would differ. New fates would ensue as the new vegetal cells activated their skeletogenic mesenchyme determinants and started activation of their neighbors. To see if this was indeed the case, Ransick and Davidson (1993) transplanted micromeres into the animal cap of an 8-cell sea urchin embryo. These skeletogenic mesenchyme precursor cells induced the formation of the archenteron and the expression of the archenteron-specific genes. The fluorescently labeled micromeres did not contribute structurally to the secondary archenteron, but provided a short-range inductive signal that respecified the animal cap cells.
Literature Cited
Davidson, E. H. 1989. Lineage-specific gene expression and the regulative capacities of the sea urchin embryo: a proposed mechanism. Development 105: 421-446.
Hough-Evans, B. R., Franks, R. R., Cameron, R. A., Britten, R. J. and Davidson, E. H. 1987. Correct cell type-specific expression of a fusion gene injected into sea urchin eggs. Dev. Biol. 121: 576-579.
Ransick, A. and Davidson, E. H. 1993. A complete second gut induced by transplanted micromeres in the sea urchin embryo. Science 259: 1134-1138.
Sucov, H. M., Hough-Evans, B. R., Franks, R. R., Britten, R. J. and Davidson, E. H. 1988. A regulatory domain that directs lineage-specific expression of a skeletal matrix protein in the sea urchin embryo. Genes Dev. 2: 1238-1250.
© All the material on this website is protected by copyright. It may not be reproduced in any form without permission from the copyright holder.
HOME :: CHAPTER 8 :: SEA URCHIN CELL SPECIFICATION :: TRANSCRIPTION FACTOR ACTIVATION IN THE SEA URCHIN EMBRYO PREVIOUS :: NEXT
Home Link
هیچ نظری موجود نیست:
ارسال یک نظر