
تخصصی شدن سین سیشیال
این نوع تخصصی شدن در حشرات دیده می شود.پس از لقاح تقسیم هسته درون سیتوپلاسم سلول تخم صورت گرفته و هسته های زیادی درون یک سلول تخم بسیار بزرگایجاد می شود. سیتوپلاسمی که شامل تعداد زیادی هسته است سین سیتیوم نامیده می شود. سیتوپلاسم سلول تخم یک پارچه نیست و ناحیه ی قدامی از ناحیه خلفی متفاوت است و بر هم کنش های تخصصی شدن سین سیشیال بین سلولها صورت نمی گیرد بلکه بین بخش های مختلف یک سلول انجام می شود.
تفاوت در ناحیه قدامی و خلفی به علت وجود اطلاعاتی است که توسط پروتئینهایی به نام ریخت زا Morphogen) ) ایجاد می شود. ریخت زاها در نواحی خاصی از جنین سنتز شده و تا فاصله زیادی منتشر می شوند. بیشترین غلظت این پروتئین ها (ریخت زاها) در نقطه سنتز است و هم زمان با دور شدن از منبع ، غلظتش کاهش می یابد. غلظت عوامل ریخت زای ویژه در هر مکان خاص به سلول می گوید که نسبت به منبع عامل ریخت زا در چه موقعیتی قرار گرفته است.
جنین شناسان نشان داده اند که در قدامی ترین بخش جنین دروزوفیلا پروتئین ریخت زایی سنتز می شود که Bicoid نامیده می شود و خلفی ترین بخش نیز سنتز کننده پروتئین Nanos می باشد. شیب غلظت این دو ماده به سمت مقابل کاهش می یابد. بنابراین محور طولی سلول تخم دروزوفیلا با دو شیب غلظت در دو سمت آن تعیین می شود. هم زمان با تقسیم هسته و ورود آنها به نواحی مختلف سیتوپلاسم سین سیتیوم ، هسته ها با توجه به نسبت Bicoid /Nanos در امتداد محور قدامی_خلفی آموزش داده می شوند. به این صورت که هسته هایی که در نواحی با غلظت بالای Bicoid و غلظت کم Nanos هستند، ژن هایی را بیان می کنند که برای تولید سر ضروری است و هسته های مستقر در نواحی با غلظت کمتر Bcoid و مقدار کمی Nanos بیان کننده ژنهایی هستند که باعث شکل گیری ناحیه سینه می شود. هم چنین هسته ها یی که در نواحی واجد میزان بسیار کم یا فاقدBicoid و غلظت بالای Nanos هستند،ساختارهای شکمی را بوجود می آورند.
شواهد مختلف توسط دانشمندان جنین شناسی نشان دهنده این موضوع است که:
1- پروتئینBicoid دارای شیبی است که بیشترین غلظت آن در ناحیه قدامی (ناحیه تشکیل سر) یافت می شود.
2- جنین های فاقد پروتئین Bicoid بدون سر هستند.
3- زمانی که Bicoid به نواحی مختلف جنین ها ی فاقد Bicoid تزریق شد، در مکان های تزریق، سر شکل گرفت. علاوه بر این در نواحی اطراف محل تزریق Bicoid با توجه به میزان غلظت Bicoid در نواحی اطراف تزریق، ناحیه ی سینه شکل گرفت( همانطور که از پیام وابسته به انتظار می رفت).
تخصصی شدن خودبخودی
انواع تخصصی شدن سلول ها در یک پست جداگانه به طور مختصر آورده شد. حال در چند پست دیگر سه نوع تخصصی شدن به طور کلی تر بیان می شود.
تخصصی شدن خود بخودی:
این روش برای اولین بار در سال 1887 توسط یک دانشجوی پزشکی فرانسوی به نام Laurent chabry مطرح شد. او روی تکوین جنین تونیکات بررسی می کرد. این جانوران در نواحی ساحلی به فراوانی یافت می شوند و دارای سلول های بزرگ هستند. جنین های تونیکات سریعا به لاروی با تعداد و تنوع کم سلولی تکوین پیدا می کند. Chabry با جداسازی بلاستومرهای جنین های در حال تقسیم تونیکات و قرار دادن آنا در محیط کشت جداگانه، دریافت که هر بلاستومر مسئول ایجاد ناحیه خاصی از بافت لارو است. در غیاب بلاستومرهای خاص، لارو فاقد همان ساختارهایی می شد که در حالت طبیعی توسط آن بلاستومرها شکل می گرفتندو هم چنین سلول های معین جداشده نیز قادر بودند فقط ساختارهای مشخصی را به وجود آورند.
بطور کلی مشخص شده که اگر از جنین 8 سلولی، سلولهای خاصی جدا شوند، جنین حاصله فاقد ساختارهای مشتق شده از آن سلول های خاص می شود درحالیکه سلول های جداشده ی دور ازجنین قادرند همان ساختارها را ایجاد کنند. به این ترتیب می توان گفت در روش تکوین خود بخودی که به آن تکوین موزائیکی نیز گفته میشود، جنین ساختاری شبیه به موزائیک با اجزای غیروابسته است که هر یک به صورت جداگانه تمایز می یابند. در جنین بی مهرگان شامل نرم تنان، کرم های حلقوی و تونیکات ها،تعیین سرنوشت سلولی اغلب توسط تخصصی شدن خودبخودی صورت می گیرد. در این موجودات، عوامل تعیین کننده ریخت زایی (شامل پروتئین ها و mRNA ) در نواحی متفاوتی از سیتوپلاسم سلول تخم قرار گرفته اند و هم زمان با تقسیمات جنینی(تسهیم) وارد سلول های مختلف شده و در نتیجه سلول های بلاستومر حاصل تخصصی شده می شوند و از هر بلاستومر دود مانی مشابه شکل میگیرد. بنابراین می توان گفت که سرنوشت بلاستومر ها ثابت است. در این روش، تخصصی شدن انواع سلولی ( بلاستومرها) قبل از هر گونه مهاجرت سلول های جنینی دیده میشود.
۱۳۸۹ دی ۱, چهارشنبه
۱۳۸۹ آذر ۷, یکشنبه
بخشی از سمینارهای ارائه شده توسط دانشجویان دکتر مینو محمودی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:Aurelia The sea moonj
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طاهره شکیبایی زاده
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:فرومون و فرومون جنسی در دروزوفیلا
استاد راهنما : دکتر مینو محمودی
گردآورندگان:زهرا عباسی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:تسهیم تخم در پرندگان
استاد راهنما : دکتر مینو محمودی
گردآورندگان:شهرزاد داروگر
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:بیان 3BMP درطی رشد اندام جوجه
استاد راهنما : دکتر مینو محمودی
گردآورندگان:فهیمه شعبانلو
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:Fish reproduction
استاد راهنما : دکتر مینو محمودی
گردآورندگان:سمیه شمس الهی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Sea Star Embryology
استاد راهنما : دکتر مینو محمودی
گردآورندگان:نسرین هاشمی فیروزی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: تعیین جنسیت در پرندگان
استاد راهنما : دکتر مینو محمودی
گردآورندگان: شهین شبرندی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Mechanisms of Branching Morphogenesis in Diverse Systems
استاد راهنما : دکتر مینو محمودی
گردآورندگان: شهین شبرندی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Basics of Molecular Cloning
استاد راهنما : دکتر مینو محمودی
گردآورندگان:محمد صادق حیرانیان
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: The Evolution of the Eye!!!
استاد راهنما : دکتر مینو محمودی
گردآورندگان: محمد شفیعی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: تکامل قلب
استاد راهنما : دکتر مینو محمودی
گردآورندگان: فاطمه عبدلله زاده
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: الگو ها(متد ها) شروع بیان ژن هموتیک
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طیبه توسن
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Flavonoids and sesquiterpenoids
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طیبه توسن
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : جنین مرغ
استاد راهنما : دکتر مینو محمودی
گردآورندگان:نرگس آرین جاه
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : گاسترولاسیون در زنوپوس
استاد راهنما : دکتر مینو محمودی
گردآورندگان:الناز نقوی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : Gastrulation in sea urchin
استاد راهنما : دکتر مینو محمودی
گردآورندگان:فاطمه عباسی تبار
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع :بررسی رفتارهای جفت گیری در پرندگان
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طلیعه شیرافکن
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع :کیست های تخمدانی
استاد راهنما : دکتر مینو محمودی
گردآورندگان:نوشین فرهمندلو
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : تکوین
استاد راهنما : دکتر مینو محمودی
گردآورندگان: فاطمه یعقوبیان
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع :گاسترولاسیون در جنین انسان
استاد راهنما : دکتر مینو محمودی
گردآورندگان: عباس قربانخانی. غلام رضا عابدی.حمیدرضا غیایی
موضوع :گاسترولاسیون در اسفنجها
استاد راهنما : دکتر مینو محمودی
گردآورندگان: غلام رضا عابدی
دانشگاه آزاد همدان
موضوع:Aurelia The sea moonj
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طاهره شکیبایی زاده
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:فرومون و فرومون جنسی در دروزوفیلا
استاد راهنما : دکتر مینو محمودی
گردآورندگان:زهرا عباسی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:تسهیم تخم در پرندگان
استاد راهنما : دکتر مینو محمودی
گردآورندگان:شهرزاد داروگر
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:بیان 3BMP درطی رشد اندام جوجه
استاد راهنما : دکتر مینو محمودی
گردآورندگان:فهیمه شعبانلو
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع:Fish reproduction
استاد راهنما : دکتر مینو محمودی
گردآورندگان:سمیه شمس الهی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Sea Star Embryology
استاد راهنما : دکتر مینو محمودی
گردآورندگان:نسرین هاشمی فیروزی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: تعیین جنسیت در پرندگان
استاد راهنما : دکتر مینو محمودی
گردآورندگان: شهین شبرندی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Mechanisms of Branching Morphogenesis in Diverse Systems
استاد راهنما : دکتر مینو محمودی
گردآورندگان: شهین شبرندی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Basics of Molecular Cloning
استاد راهنما : دکتر مینو محمودی
گردآورندگان:محمد صادق حیرانیان
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: The Evolution of the Eye!!!
استاد راهنما : دکتر مینو محمودی
گردآورندگان: محمد شفیعی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: تکامل قلب
استاد راهنما : دکتر مینو محمودی
گردآورندگان: فاطمه عبدلله زاده
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: الگو ها(متد ها) شروع بیان ژن هموتیک
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طیبه توسن
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع: Flavonoids and sesquiterpenoids
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طیبه توسن
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : جنین مرغ
استاد راهنما : دکتر مینو محمودی
گردآورندگان:نرگس آرین جاه
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : گاسترولاسیون در زنوپوس
استاد راهنما : دکتر مینو محمودی
گردآورندگان:الناز نقوی
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : Gastrulation in sea urchin
استاد راهنما : دکتر مینو محمودی
گردآورندگان:فاطمه عباسی تبار
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع :بررسی رفتارهای جفت گیری در پرندگان
استاد راهنما : دکتر مینو محمودی
گردآورندگان:طلیعه شیرافکن
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع :کیست های تخمدانی
استاد راهنما : دکتر مینو محمودی
گردآورندگان:نوشین فرهمندلو
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع : تکوین
استاد راهنما : دکتر مینو محمودی
گردآورندگان: فاطمه یعقوبیان
مجموعه کنفرانسهای علمی انجمن علمی
دانشگاه آزاد همدان
موضوع :گاسترولاسیون در جنین انسان
استاد راهنما : دکتر مینو محمودی
گردآورندگان: عباس قربانخانی. غلام رضا عابدی.حمیدرضا غیایی
موضوع :گاسترولاسیون در اسفنجها
استاد راهنما : دکتر مینو محمودی
گردآورندگان: غلام رضا عابدی
۱۳۸۹ آبان ۳۰, یکشنبه
سمینار خانم نادری.استاد مربوطه:دکتر محمودی
به نام پروردگاری که هنوز ذهن بشر در برابر آفریده هایش درمانده است.
Origin, Development and Regulation of Human Leydig Cells
Azam Naderi Farjam
Hamedan Islamic Azad University
Advisor: Dr.M.mahmoodi
2o10
1-Stem Leydig Cell : 2-Progenitor Leydig Cell : 3-Immature Leydig Cell :
4-Mature (Adult) Leydig Cell:
The development and maturation of Leydig cells are dynamic processes involving interaction between hormones and numerous additional factors. In humans, fetal and adult populations of Leydig cells with distinct lineages have been described. During the embryonic and fetal period, these cells secrete testosterone and other androgens, which regulate not only the masculinization of internal and external genitalia,
The development of fetal Leydig cells in humans is a complex process involving cascades of cellular events leading to proliferation, differentiation and involution and associated specific changes in morphology and function. The close functional similarity between fetal Leydig cells and adrenocortical cells found recently suggests a common origin during embryogenesis.. These cells possess well-developed steroidogenic machinery expressing both the luteinizing hormone receptor (LHR) (fig. 1a) and the key steroidogenic enzymes (e.g. 3βHSD, P450scc, P450c17) required for androgen biosynthesis
Immunohistochemical analysis of the expression of receptors and steroidogenic enzymes by Leydig cells and their putative precursors isolated from the rat testis. a Expression of LH receptor by fetal Leydig cells isolated from 7-day-old rats. b Expression of PDGF receptor-α by putative Leydig stem cells isolated from rats 7 days postnatally. c Expression of the steroidogenic acute regulatory (StAR) protein by immature Leydig cells. d Expression of cytochrome P450scc by adult-type Leydig cells. e Expression of α-actin by peritubular cells isolated from 20-day-old rats.
The neonatal androgen surge (‘baby puberty’) may play a role in imprinting various cell types in the prostate, kidney and brain in such a manner they respond appropriately to androgen stimulation during adulthood. the Leydig cells are stimulated by the pituitary gonadotropin LH to grow in number and cellular size, and, the pubertal surge of testosterone required for start and maintenance of full spermatogenesis, development of the accessorysex glands and the appearance of the secondary sexual characteristics.
The development of fetal Leydig cells can be separated into three stages, i.e. differentiation, fetal maturation and involution In the embryonic and fetal human testis, differentiation occurs at a gestational age of 7–14 weeks; maturation during the weeks 14–18 of gestation; and the involution thereafter until the time of full-term birth. The maximal number of Leydig cells per pair of human testes (48 × 106) is observed ig Cells during weeks 13–16 of gestation The Leydig cells appear embryonic gonad shortly after testis determination and probably arise from multiple embryonic tissues including the coelomic epithelium, gonadal ridge mesenchyme, and migrating mesonephric cells .
Moreover, it has been proposed that fetal Leydig cells may have initially evolved through slight modifications of the fetal adrenal cells. This suggestion is based on the observation that patients suffering from congenital adrenal hyperplasia, which in inadequately controlled cases may be associated with chronically elevated levels of ACTH, develop testicular masses known as adrenal rest tissue [8]. This tissue is generally thought to arise from ectopic adrenal tissue which has failed to separate from the gonad during fetal differentiation .However, it is also possible that in some cases these masses arise from ACTH-sensitive fetal Leydig cells that are probably still present in the testis. Of interest in this context is the observation that chronically elevated secretion of ACTH in a boy to stimulate androgen production and cause precocious puberty .
The precursors of fetal human Leydig cells become functionally active as early as after 6–7 weeks of gestation, at which time testosterone can be detected in the human embryonic testis. The differentiation of these cells must be independent of LH, since the onset of testicular androgen production precedes the secretion of LH by the fetal pituitary. Recently, Lambrot et al. have added more support to this conclusion demonstrating that the capacity of the embryonic testis to produce ., retinoic acid can stimulate the expression of the key steroidogenic enzymes and, thereby, steroidogenesis by human embryonic Leydig cells . These observations provide strong support for the idea that neither hCG secreted by the placenta nor LH are involved in regulating the initial phase of human Leydig cell differentiation. However, LH does stimulate testicular testosterone production after more than 7 weeks of gestation, indicating that hCG/LH is absolutely required for the maintenance of this production at later
Moreover, gonadotropin insufficiency is often associated with undermasculinization including micropenis, which is an indication of insufficient testosterone production by fetal Leydig cells . Similarly, males carrying an inactivating mutation of the LHR exhibit reduced numbers of Leydig cells and show poor development of the external genitalia . However, a certain development of androgen-sensitive organs (e.g. the ductus deferens and epididymis) may occur in these individuals indicating some LH/hCG-independent production of androgen,., mutation of the LHβ gene that eliminates the ability of this hormone to bind to its receptor eliminated Leydig cell development in a patient with male hypogonadism.
The Leydig cells proliferate and differentiate gradually and continuously until they attain peak development and maturation before week 19 of gestation, followed by regression .To date, there is no information available concerning the testicular and/or circulating factor(s) that triggers the degeneration of fetal human Leydig cells. Interestingly, in rodents this regression occurs when plasma levels of LH are still high , indicating that this gonadotropin is unable to protect the cells from involution. Several signal molecules, including transforming growth factor (TGF)-β, anti-Müllerian hormone (AMH) and gonadotropin-releasing hormone (GnRH), have been proposed to play a role in the degeneration of fetal Leydig cells in rodents .
Regulatory Factors Controlling the Functions of Fetal HumaHuman Leydig Cells
Desert Hedgehog Although most data concerning the role of Desert hedgehog (Dhh) in the regulation of fetal Leydig cells have been obtained in rodents, there are also reports on the significant role of Dhh signaling in the development normal testicular phenotype in humans . In rodents, Dhh is known to be required for the differentiation and expansion of fetal Leydig cells during the embryonic phase. Secreted by the Sertoli cell, Dhh acts in a paracrine fashion to induce the differentiation of both the fetal Leydig cells and of the peritubular myoid cells, surrounding the testicular cords . Mutant mouse male gonads that lack functional Dhh contain no Leydig cells and the consequent absence of androgens results in feminization of the external genitalia of these animals. Recently compound Sf1+/– ; Dhh –/– mutant male mice were shown not to masculinize and to appear externally female as a result of their lack of differentiated fetal Leydig cells. These findings led to the proposal that the Sf1 and Dhh pathways may be necessary for the differentiation and survival of fetal and adult Leydig cells in rodent Dhh in humans still remains to be demonstrated.
Human fetal Leydig cells have been found to express significant levels of growth factor of both isoforms, PDGF-A and PDGF-B, as well as their corresponding receptors, PDGFRα and PDGFRβ . Therefore, it is reasonable to suggest that the PDGF system is involved in the control of Leydig cell development and function also in humans, similarly to what has been observed in rodents. Deletion of the gene encoding PDGF-A was found to attenuate the expression of cytochrome P450 side-chain cleavage (P450scc) , which converts cholesterol into pregnenolone, and thereby disrupts early Leydig cell I differentiation in mice.
The expression of this regulatory molecule in human fetal Leydig cells has recently been shown to peak week 15 postconception, in parallel with the highest testosterone levels of the fetus , suggesting a role for GATA-4 in the regulation of steroidogenesis in human fetal Leydig cell. Experiments on rodents have shown that GATA-4 plays a role in the differentiation of and/or steroidogenesis by somatic gonadal cells, including fetal and adult Leydig cells,
Furthermore, some evidence suggests that ligands of the IGF system affect the differentiation and steroidogenic capacity of fetal Leydig cells, both in rodents and humans . In the human testis, IGFs and the type I IGF receptor are differentially expressed at different maturational stages and seem to be involved in the regulation of Leydig cell proliferation and survival, and steroidogenic maturation . In line with this hypothesis, boys with a genetic deficiency in production of this growth factor are undermasculinized,
Postnatal Differentiation of Human Leydig Cells
Division of the development of human Leydig cells into the three stages is based on the triphasic development of plasma testosterone levels, with the initial testosterone peak appearing at the end of the first trimester of fetal life, when mature fetal Leydig cells are functional, the second after 2–3 months of postnatal life, and the third peak being established in connection with established puberty and lasting thereafter throughout adulthood, until a decline at old age. Morphometric analysis reflects this fluctuation in testosterone levels, revealing a decline in the number of fetal Leydig cells following their peak development during 14–18 weeks of gestation and the appearance of a new wave of Leydig cells designated as neonatal 2–3 months after birth. In humans, the development and function of neonatal Leydig cells are thought to be controlled by the elevated level of LH, which originates from reactivation of the hypothalamic-pituitary-gonadal (HPG) axis during the neonatal period , and is associated with an increased plasma testosterone level. Thus, blockage of the neonatal activation of the HPG axis by an antagonist of GnRH suppresses the maturation of Leydig cells in primates and abolishes the neonatal rise in testosterone secretion .
The enhancement in the numbers of mature Leydig cells that occurs during puberty indicates that precursor cells (e.g. peritubular-like Leydig stem cells and perivascular cells) are being recruited for this purpose . The peritubular-like Leydig stem cells express PDGFRα (fig. 1b) but not the LHR or steroidogenic enzymes . After experimental ablation of Leydig cells with ethane dimethane sulfonate (EDS), peritubular spindle-shaped cells have been shown to be precursors of regenerating Leydig cells in rats . Interestingly, certain investigations on both humans and experimental animals have demonstrated that fully mature Leydig cells can dedifferentiate back to their previous stage of development. This event is associated with several morphological changes, including reductions in the volume of the smooth endoplasmic reticulum and number of mitochondria, as well as with impairment of testosterone secretion
Fig. 2. A putative scheme for the differentiation of human Leydig cells. Peritubular (PTC) and Sertoli cells (SC) secrete a number of critical factors including LIF, PDGF-α and Dhh which trigger Leydig stem cells (LSC) to proliferate and migrate into the interstitial compartment of the testis, where they differentiate into progenitor Leydig cells (PLC). Subsequently, a combination of growth factors and hormones (e.g. LH, T3, IGF-1 and PDGF-α) activate their signaling pathways that promote transition of the PLC into immature Leydig cells (ILC) and, finally, into the adult Leydig cell (ALC) population.
Regulation of the Function of Human Adult Leydig Cell Lineage
Insulin-Like Factor 3 Insulin-like factor 3 (INSL3), a peptide that controls the early phase of testicular descent during embryonic development, is expressed and secreted not only by fetal but also by adult-type human Leydig cells. Importantly. Recently, the INSL3 concentration in the peripheral blood of men has been shown to decline continuously in a linear fashion between the ages of 35 and 80, a phenomenon that probably reflects a reduction in Leydig cell functionality with age All these reports agree well with recent findings showing that testosterone upregulates the expression of INSL3 in primary rat Leydig cell cultures suggesting a positive correlation between testosterone production and INSL3 expression by Leydig cells.
Ghrelin The peptide ghrelin, an endogenous ligand for growth hormone, detected in the human stomach, hypothalamus and testis, is also an important regulator of human Leydig cell function. In the case of the testis, ghrelin has been localized immunohistochemically to Leydig cells and Sertoli cells.
This peptide inhibits both hCG- and cAMP-activated testosterone production by Leydig cells in vitro, an effect which is associated with significant attenuation of hCG-stimulated expression of mRNAs coding for several key steroidogenic proteins, e.g. StAR, P450scc, 3β-HSD, and 17β-HSD type III . Recently, the level of ghrelin expression by Leydig cells was found to be inversely correlated to serum levels of testosterone in patients ,in seminiferous tubules of the rat testis ghrelin regulates the expression of the stem cell factor (SCF), a major paracrine stimulator of germ cell development which promotes the survival of spermatogonia, spermatocytes, and spermatids in the adult rat seminiferous epithelium.
Leptin Leptin, a 16-kDa protein produced primarily by adipose tissue [68] and exerting a significant influence on reproduction and fertility in mammals. Experiments on rodents have shown that leptin is a potent suppressor of steroidogenesis in Leydig cells. The rapid and dose-dependent inhibition of hCG-stimulated testosterone production by primary cultures of rat Leydig cells exerted by leptin is associated with attenuation of androstenedione levels and a concomitant elevation of the levels of the precursor molecules 17-OH progesterone, progesterone and pregnenolone, Moreover, leptin reduces the expression of mRNAs encoding SF-1, StAR and P450scc, which are key elements of the steroidogenic machinery in this same system . These effects in rodent models are consistent with those observed in humans. Obese individuals demonstrate elevated levels of leptin and reduced concentrations of androgens in their blood [72]. Furthermore, circulating concentrations of leptin have been suggested to be the most reliable predictor of obesity-related attenuation of the androgen response to hCG in vitro. Recently, leptin was demonstrated to be expressed in germ cells, while the expression of the leptin receptor was found exclusively in the Leydig cells of fertile men ,leptin receptor expression in Leydig cells is inversely correlated with the serum levels of testosterone. Thus, overexpression of the leptin receptor by Leydig cells appears to inhibit testosterone production in infertile men.
At present, our knowledge concerning the intracellular signaling cascades that control the development of different populations of human Leydig cells is still incomplete.
In addition, possible reduction of the capacity of fetal human Leydig cells to produce androgens induced by xenobiotics (see, for example ), which may lead to incomplete masculinization of male fetuses and various malformations in the reproductive tract, should be explored.
The signaling molecules that trigger the dedifferentiation of fully mature Leydig cells to their previous stage of development are completely unknown and a better understanding of this phenomenon may provide insight into the mechanisms underlying aging and tumorigenesis.
Figure 1 is a scanning electron micrograph of a cross-section of the rat testis . The micrograph shows the beautiful architecture of the testis. The seminiferous tubules are quite prominent and reveals tubules at different stages of spermatogenesis. The web like interstitium that surrounds the tubules is where Leydig cells, testicular macrophages, endothelial cells and other interstitial cells reside.
Figure 3: testosterone biosynthetic pathway in Leydig cells.
the end
Origin, Development and Regulation of Human Leydig Cells
Azam Naderi Farjam
Hamedan Islamic Azad University
Advisor: Dr.M.mahmoodi
2o10
1-Stem Leydig Cell : 2-Progenitor Leydig Cell : 3-Immature Leydig Cell :
4-Mature (Adult) Leydig Cell:
The development and maturation of Leydig cells are dynamic processes involving interaction between hormones and numerous additional factors. In humans, fetal and adult populations of Leydig cells with distinct lineages have been described. During the embryonic and fetal period, these cells secrete testosterone and other androgens, which regulate not only the masculinization of internal and external genitalia,
The development of fetal Leydig cells in humans is a complex process involving cascades of cellular events leading to proliferation, differentiation and involution and associated specific changes in morphology and function. The close functional similarity between fetal Leydig cells and adrenocortical cells found recently suggests a common origin during embryogenesis.. These cells possess well-developed steroidogenic machinery expressing both the luteinizing hormone receptor (LHR) (fig. 1a) and the key steroidogenic enzymes (e.g. 3βHSD, P450scc, P450c17) required for androgen biosynthesis
Immunohistochemical analysis of the expression of receptors and steroidogenic enzymes by Leydig cells and their putative precursors isolated from the rat testis. a Expression of LH receptor by fetal Leydig cells isolated from 7-day-old rats. b Expression of PDGF receptor-α by putative Leydig stem cells isolated from rats 7 days postnatally. c Expression of the steroidogenic acute regulatory (StAR) protein by immature Leydig cells. d Expression of cytochrome P450scc by adult-type Leydig cells. e Expression of α-actin by peritubular cells isolated from 20-day-old rats.
The neonatal androgen surge (‘baby puberty’) may play a role in imprinting various cell types in the prostate, kidney and brain in such a manner they respond appropriately to androgen stimulation during adulthood. the Leydig cells are stimulated by the pituitary gonadotropin LH to grow in number and cellular size, and, the pubertal surge of testosterone required for start and maintenance of full spermatogenesis, development of the accessorysex glands and the appearance of the secondary sexual characteristics.
The development of fetal Leydig cells can be separated into three stages, i.e. differentiation, fetal maturation and involution In the embryonic and fetal human testis, differentiation occurs at a gestational age of 7–14 weeks; maturation during the weeks 14–18 of gestation; and the involution thereafter until the time of full-term birth. The maximal number of Leydig cells per pair of human testes (48 × 106) is observed ig Cells during weeks 13–16 of gestation The Leydig cells appear embryonic gonad shortly after testis determination and probably arise from multiple embryonic tissues including the coelomic epithelium, gonadal ridge mesenchyme, and migrating mesonephric cells .
Moreover, it has been proposed that fetal Leydig cells may have initially evolved through slight modifications of the fetal adrenal cells. This suggestion is based on the observation that patients suffering from congenital adrenal hyperplasia, which in inadequately controlled cases may be associated with chronically elevated levels of ACTH, develop testicular masses known as adrenal rest tissue [8]. This tissue is generally thought to arise from ectopic adrenal tissue which has failed to separate from the gonad during fetal differentiation .However, it is also possible that in some cases these masses arise from ACTH-sensitive fetal Leydig cells that are probably still present in the testis. Of interest in this context is the observation that chronically elevated secretion of ACTH in a boy to stimulate androgen production and cause precocious puberty .
The precursors of fetal human Leydig cells become functionally active as early as after 6–7 weeks of gestation, at which time testosterone can be detected in the human embryonic testis. The differentiation of these cells must be independent of LH, since the onset of testicular androgen production precedes the secretion of LH by the fetal pituitary. Recently, Lambrot et al. have added more support to this conclusion demonstrating that the capacity of the embryonic testis to produce ., retinoic acid can stimulate the expression of the key steroidogenic enzymes and, thereby, steroidogenesis by human embryonic Leydig cells . These observations provide strong support for the idea that neither hCG secreted by the placenta nor LH are involved in regulating the initial phase of human Leydig cell differentiation. However, LH does stimulate testicular testosterone production after more than 7 weeks of gestation, indicating that hCG/LH is absolutely required for the maintenance of this production at later
Moreover, gonadotropin insufficiency is often associated with undermasculinization including micropenis, which is an indication of insufficient testosterone production by fetal Leydig cells . Similarly, males carrying an inactivating mutation of the LHR exhibit reduced numbers of Leydig cells and show poor development of the external genitalia . However, a certain development of androgen-sensitive organs (e.g. the ductus deferens and epididymis) may occur in these individuals indicating some LH/hCG-independent production of androgen,., mutation of the LHβ gene that eliminates the ability of this hormone to bind to its receptor eliminated Leydig cell development in a patient with male hypogonadism.
The Leydig cells proliferate and differentiate gradually and continuously until they attain peak development and maturation before week 19 of gestation, followed by regression .To date, there is no information available concerning the testicular and/or circulating factor(s) that triggers the degeneration of fetal human Leydig cells. Interestingly, in rodents this regression occurs when plasma levels of LH are still high , indicating that this gonadotropin is unable to protect the cells from involution. Several signal molecules, including transforming growth factor (TGF)-β, anti-Müllerian hormone (AMH) and gonadotropin-releasing hormone (GnRH), have been proposed to play a role in the degeneration of fetal Leydig cells in rodents .
Regulatory Factors Controlling the Functions of Fetal HumaHuman Leydig Cells
Desert Hedgehog Although most data concerning the role of Desert hedgehog (Dhh) in the regulation of fetal Leydig cells have been obtained in rodents, there are also reports on the significant role of Dhh signaling in the development normal testicular phenotype in humans . In rodents, Dhh is known to be required for the differentiation and expansion of fetal Leydig cells during the embryonic phase. Secreted by the Sertoli cell, Dhh acts in a paracrine fashion to induce the differentiation of both the fetal Leydig cells and of the peritubular myoid cells, surrounding the testicular cords . Mutant mouse male gonads that lack functional Dhh contain no Leydig cells and the consequent absence of androgens results in feminization of the external genitalia of these animals. Recently compound Sf1+/– ; Dhh –/– mutant male mice were shown not to masculinize and to appear externally female as a result of their lack of differentiated fetal Leydig cells. These findings led to the proposal that the Sf1 and Dhh pathways may be necessary for the differentiation and survival of fetal and adult Leydig cells in rodent Dhh in humans still remains to be demonstrated.
Human fetal Leydig cells have been found to express significant levels of growth factor of both isoforms, PDGF-A and PDGF-B, as well as their corresponding receptors, PDGFRα and PDGFRβ . Therefore, it is reasonable to suggest that the PDGF system is involved in the control of Leydig cell development and function also in humans, similarly to what has been observed in rodents. Deletion of the gene encoding PDGF-A was found to attenuate the expression of cytochrome P450 side-chain cleavage (P450scc) , which converts cholesterol into pregnenolone, and thereby disrupts early Leydig cell I differentiation in mice.
The expression of this regulatory molecule in human fetal Leydig cells has recently been shown to peak week 15 postconception, in parallel with the highest testosterone levels of the fetus , suggesting a role for GATA-4 in the regulation of steroidogenesis in human fetal Leydig cell. Experiments on rodents have shown that GATA-4 plays a role in the differentiation of and/or steroidogenesis by somatic gonadal cells, including fetal and adult Leydig cells,
Furthermore, some evidence suggests that ligands of the IGF system affect the differentiation and steroidogenic capacity of fetal Leydig cells, both in rodents and humans . In the human testis, IGFs and the type I IGF receptor are differentially expressed at different maturational stages and seem to be involved in the regulation of Leydig cell proliferation and survival, and steroidogenic maturation . In line with this hypothesis, boys with a genetic deficiency in production of this growth factor are undermasculinized,
Postnatal Differentiation of Human Leydig Cells
Division of the development of human Leydig cells into the three stages is based on the triphasic development of plasma testosterone levels, with the initial testosterone peak appearing at the end of the first trimester of fetal life, when mature fetal Leydig cells are functional, the second after 2–3 months of postnatal life, and the third peak being established in connection with established puberty and lasting thereafter throughout adulthood, until a decline at old age. Morphometric analysis reflects this fluctuation in testosterone levels, revealing a decline in the number of fetal Leydig cells following their peak development during 14–18 weeks of gestation and the appearance of a new wave of Leydig cells designated as neonatal 2–3 months after birth. In humans, the development and function of neonatal Leydig cells are thought to be controlled by the elevated level of LH, which originates from reactivation of the hypothalamic-pituitary-gonadal (HPG) axis during the neonatal period , and is associated with an increased plasma testosterone level. Thus, blockage of the neonatal activation of the HPG axis by an antagonist of GnRH suppresses the maturation of Leydig cells in primates and abolishes the neonatal rise in testosterone secretion .
The enhancement in the numbers of mature Leydig cells that occurs during puberty indicates that precursor cells (e.g. peritubular-like Leydig stem cells and perivascular cells) are being recruited for this purpose . The peritubular-like Leydig stem cells express PDGFRα (fig. 1b) but not the LHR or steroidogenic enzymes . After experimental ablation of Leydig cells with ethane dimethane sulfonate (EDS), peritubular spindle-shaped cells have been shown to be precursors of regenerating Leydig cells in rats . Interestingly, certain investigations on both humans and experimental animals have demonstrated that fully mature Leydig cells can dedifferentiate back to their previous stage of development. This event is associated with several morphological changes, including reductions in the volume of the smooth endoplasmic reticulum and number of mitochondria, as well as with impairment of testosterone secretion
Fig. 2. A putative scheme for the differentiation of human Leydig cells. Peritubular (PTC) and Sertoli cells (SC) secrete a number of critical factors including LIF, PDGF-α and Dhh which trigger Leydig stem cells (LSC) to proliferate and migrate into the interstitial compartment of the testis, where they differentiate into progenitor Leydig cells (PLC). Subsequently, a combination of growth factors and hormones (e.g. LH, T3, IGF-1 and PDGF-α) activate their signaling pathways that promote transition of the PLC into immature Leydig cells (ILC) and, finally, into the adult Leydig cell (ALC) population.
Regulation of the Function of Human Adult Leydig Cell Lineage
Insulin-Like Factor 3 Insulin-like factor 3 (INSL3), a peptide that controls the early phase of testicular descent during embryonic development, is expressed and secreted not only by fetal but also by adult-type human Leydig cells. Importantly. Recently, the INSL3 concentration in the peripheral blood of men has been shown to decline continuously in a linear fashion between the ages of 35 and 80, a phenomenon that probably reflects a reduction in Leydig cell functionality with age All these reports agree well with recent findings showing that testosterone upregulates the expression of INSL3 in primary rat Leydig cell cultures suggesting a positive correlation between testosterone production and INSL3 expression by Leydig cells.
Ghrelin The peptide ghrelin, an endogenous ligand for growth hormone, detected in the human stomach, hypothalamus and testis, is also an important regulator of human Leydig cell function. In the case of the testis, ghrelin has been localized immunohistochemically to Leydig cells and Sertoli cells.
This peptide inhibits both hCG- and cAMP-activated testosterone production by Leydig cells in vitro, an effect which is associated with significant attenuation of hCG-stimulated expression of mRNAs coding for several key steroidogenic proteins, e.g. StAR, P450scc, 3β-HSD, and 17β-HSD type III . Recently, the level of ghrelin expression by Leydig cells was found to be inversely correlated to serum levels of testosterone in patients ,in seminiferous tubules of the rat testis ghrelin regulates the expression of the stem cell factor (SCF), a major paracrine stimulator of germ cell development which promotes the survival of spermatogonia, spermatocytes, and spermatids in the adult rat seminiferous epithelium.
Leptin Leptin, a 16-kDa protein produced primarily by adipose tissue [68] and exerting a significant influence on reproduction and fertility in mammals. Experiments on rodents have shown that leptin is a potent suppressor of steroidogenesis in Leydig cells. The rapid and dose-dependent inhibition of hCG-stimulated testosterone production by primary cultures of rat Leydig cells exerted by leptin is associated with attenuation of androstenedione levels and a concomitant elevation of the levels of the precursor molecules 17-OH progesterone, progesterone and pregnenolone, Moreover, leptin reduces the expression of mRNAs encoding SF-1, StAR and P450scc, which are key elements of the steroidogenic machinery in this same system . These effects in rodent models are consistent with those observed in humans. Obese individuals demonstrate elevated levels of leptin and reduced concentrations of androgens in their blood [72]. Furthermore, circulating concentrations of leptin have been suggested to be the most reliable predictor of obesity-related attenuation of the androgen response to hCG in vitro. Recently, leptin was demonstrated to be expressed in germ cells, while the expression of the leptin receptor was found exclusively in the Leydig cells of fertile men ,leptin receptor expression in Leydig cells is inversely correlated with the serum levels of testosterone. Thus, overexpression of the leptin receptor by Leydig cells appears to inhibit testosterone production in infertile men.
At present, our knowledge concerning the intracellular signaling cascades that control the development of different populations of human Leydig cells is still incomplete.
In addition, possible reduction of the capacity of fetal human Leydig cells to produce androgens induced by xenobiotics (see, for example ), which may lead to incomplete masculinization of male fetuses and various malformations in the reproductive tract, should be explored.
The signaling molecules that trigger the dedifferentiation of fully mature Leydig cells to their previous stage of development are completely unknown and a better understanding of this phenomenon may provide insight into the mechanisms underlying aging and tumorigenesis.
Figure 1 is a scanning electron micrograph of a cross-section of the rat testis . The micrograph shows the beautiful architecture of the testis. The seminiferous tubules are quite prominent and reveals tubules at different stages of spermatogenesis. The web like interstitium that surrounds the tubules is where Leydig cells, testicular macrophages, endothelial cells and other interstitial cells reside.
Figure 3: testosterone biosynthetic pathway in Leydig cells.
the end
سمینار درس جنین شناسی تکوینی دکتر مینو محمودی
شناسایی مراحل نمو در ستاره دریایی
تسهیم
اولین مرحله ساده که تعدادی از سلولها حاوی اند. مراحل 2 سلولی, 4سلولی, 8 سلولی و الی آخر. این مراحل از اولین , دومین, سومین وتا تسهیمات پایانی حاصل می شوند. سلولهای حاصل از هر مرحله به نام بلاستومر خوانده می شوند. سلولها به سادگی بر تعدادشان افزوده شده تا مراحل 32 سلولی یا 64 سلولی , در این مراحل به جنین مورولا اطلاق می شود. مورولا توده کروی شکل حاصل از 12 تا چند صد سلول است.
بلاستولاسیون
مورولا توده توپر است ولی تسهیم ادامه یافته تا بلاستومر به سمت پیرامون خود کشیده شوند و حفره داخلی پر شده از مایع ایجاد شود. جنین یک توده از سلول هایی است که طویل نیستند ولی تشکیل یک کره را داده اند. در این مرحله جنین بلاستولا خوانده می شود و حفره پر از مایع بلاستوسل خوانده می شود. این تغییرات در حدود 10ساعت پس از لقاح به طول می انجامد. زمانی که بلاستولا برای نخست شکل می گیرد حاوی تقریبا 1000 سلول است.
گاسترولاسیون (لایه زایی)
چند ساعت کوتاه پس از بلاستولا فرایند های گاسترولاسیون (لایه زایی) آغاز می شود. در یک نقطه قطعی از سطح بلاستولا, سلولها شروع به حرکت به سمت مرکز بلاستوسل می کند. در آغاز این شکل گیری منجر به خمیدگی لوله می شود. اگر شما بادکنکی را به آرامی از آب یا هوا پر کنید و با انگشتانتان به داخل بادکنک فشار بیاوریدشما بطور تقریبی گاسرولاسیون را انجام داده اید. سطح باز شدن جنین منفذ بلاستوپور خوانده می شود و در دوتروستومیان ها (دو دهانه ای ها) نظیر توتیا و ستاره دریایی منفذ بلاستوپور یه مخرج آینده موجود تبدیل می شود. لوله خمیده آرکانترون نامیده می شود ودر نهایت به تمام جنین رسیده و در جهت دیگری از سطح جنین باز می شود و اساس شکل گیری لوله گوارشی جانور خواهد شد. از زمانی که منفذ بلاستوپور و ارکانترون شکل گیرد, جنین گاسترولا خوانده می شود.
گاسترولاسیون در جنین میکرولسیتال
در تصاویر ارایه شده، جنین در مرحله گاسترولای اولیه تا مرحله لارو" بی پی ناریا"دیده می شود.روند گاسترولاسیون مانند اندازه و توزیع زرده و تسهیم در بین گونه های مختلف در جزییات متفاوت است. در این مرحله برخی از سلول های سطح جنین به داخل حرکت کرده و لایه های سلولی جدید شکل می گیرد. این حرکات با تمایز سلول های مهاجر بداخل لایه های منفرد بافتی توام است.(ژن های خاصی فعالیت های تمایزی را باعث می شوند.)
در طی لایه زایی یک بلاستولا با خفره بلاستوسل نسبتا بزرگ شکل می گیردکه فضای کافی برای حرکت برخی از سلول های سطحی بداخل جهت تشکیل لایه های جدید را دارد.مشاهدات اولیه ستاره دریای جنین در قطب گیاهی مسطح می شود .(مهاجرت اولیه و تمایز سلول ها) که بداخل بلاستوسل قرورفتگی می یابند. و لایه سلولی داخلی شکل گرفته (هایپوبلاست یا مزودرم )و لایه های مزودرم و اندودرم شکل می گیرند.سلول های سطح خارجی گاسترولا ، اپی بلاست یا اکتودرم می شوند. و حفره بلاستوسل ، گاستروسل یاارکانترون (لوله گوارشی اولیه ) نامیده می شود. که در قطب گیاهی بعنوان بلاستوپور باز می شود.
انتهای داخلی هایپوبلاست (مزودرم) کیسه های مزودرمال را شکل می دهد. این کیسه ها Entrocelic نامیده می شوند. و اعمال انها فعالیت سلومی خوانده می شود. به موازات رشد جنین طی گاسترولاسیون ، سلول هایی از هایپوبلاست جوانه زده و برای تشکیل سلول های مزانشیمی به داخل مهاجرت میکنند.
تمایز
در مرحله گاسترولا , اندازه قطعی از تمایز نمو می یابد. سلول های سطح بیرونی گاسترولا اکتو درم را تشکیل می دهد و در آنها اپیدرم و سیستم عصبی جانور بالغ تمایز می یابد. این دولایه بوسیله لایه سوم ، مزودرم به همدیگر متصل می شوند.
در توتیا و ستاره دریایی رشد ،تمایز و شکل گیری در گاسترولا ادامه می یابد. مرحله لارو آزاد که بطور کامل قادر به حرکت و تغذیه از محیط اطراف خود است، شکل میگیرد. در مرحله بعد یک دگردیسی رخ داده و لارو به ستاره دریایی بالغ یا توتیای بالغ تبدیل می شود
تصاویری از تسهیم و لایه زایی ستاره دریایی
این صفحه یکسری از تصاویری که از تخم میکرو لیستال از سلول لقاح نیافته با مرحله بلاستولادر جنین ستاره دریایی را نشان می دهد. تخم های میکرو لیستال دارای اندازه خیلی کوچکی از زرده ذخیره شده اند. در نتیجه وجود اندازه کوچک زرده ,این حیوانات یکی از دو روش نمو را نشان میدهد.
در اولین روش لارو در مرحله جنینی از مواد تغذیه ای محیط تغذیه می شود. (در جنین های آورده شده زیر) در روش دوم که نمو داخلی است. نیاز های اساسی به وسیله مادر برآورده می شود(چیزی که ما در پستانداران مشاهده می کنیم.)
Later
تخم لقاح نیافته که از طریق اندازه و هسته مشخص (پیکان بزرگ) با هستک مشخص (پیکان کوچک) و فقدان غشای لقاحی قابل تشخیص است. تصویر زیگوت یا تخم لقاح که بوسیله غشای لقاحی (پیکانها) احاطه کرده سلول و فضای دور ویتیلینی که با مایع پر شده است،مشخص است.
Later
مرحله دو سلولی که زیگوت اولین تسهیم را کامل کرده است و تسهیم اول کامل و مساوی است.(نظیر تخمک کامل که به دو سلول تقسیم می شود). تسهیم از حول نباتی-جانوری عبور کرده است و دو بلاستومر مشابه تولید می کند. مرحله چهار سلولی که تسهیم دوم معمولا از سراسر محور گیاهی-نباتی عبور می کند. ولی بر تسهیم اول عمود است. در پایان چهار بلاستومر کامل با اندازه مساوی ایجاد می شود.
Later
مرحله هشت سلولی, سومین تسهیم در سطح استوایی رخ میدهد.(عمود بر دو تسهیم اول و محور نباتی-جانوری). چهار بلاستومر فوقانی (قطب جانوری) در اندازه کوچکتر از چهار بلاستومر تحتانی (قطب گیاهی) هستند. مرحله 32 سلولی: پس از مرحله 16 سلولی تقسیم بطور پیچیده دنبال می گردد. افزایش تعداد سلولها و تقسیمات بلاستومرها همزمان انجام می شود. تقسیم ادامه یافته و یک توده سلولی که خودش را به بلاستولا سازماندهی می کند, شکل می گیرد.
Later
بلاستولای اولیه: تسهیم ادامه پیدا می کند. در مرکز, تماس سلول ها با یکدیگر از بین رفته, و حفره مرکز پوشیده از مایع (بلاستوسل) شکل می گیرد. این بلاستوسل توسط یک لایه از سلول احاطه می شود. کره توخالی شکل گرفته به عنوان بلاستولا شناخته می شود. بلاستولای تاخیری: مشابه بلاستولای اولیه توسط یک لایه منفرد از سلول های احاطه کننده حفره مرکزی قابل تشخیص است. بلاستومرها کوچک و بطور مشخص واضح نیستند.بلاستومرهای قطب گیاهی vp ضخیم تر از قطب جانوری APظاهر می شود.
Later
گاسترولای اولیه
animal pole (AP)
vegetal pole (VP)
Later
گاسترولای میانی گاسترولای تاخیری
C
O
L
O
U
R
I
Z
E
!!
لارو بی پی ناریا
1. Blastocoel
2. Enterocoelic pouch
3. Mouth
4. Archenteron
5. Blastopore (anus) • blue=Ectoderm
• red=Mesoderm
• yellow=Endoderm
.
تسهیم
اولین مرحله ساده که تعدادی از سلولها حاوی اند. مراحل 2 سلولی, 4سلولی, 8 سلولی و الی آخر. این مراحل از اولین , دومین, سومین وتا تسهیمات پایانی حاصل می شوند. سلولهای حاصل از هر مرحله به نام بلاستومر خوانده می شوند. سلولها به سادگی بر تعدادشان افزوده شده تا مراحل 32 سلولی یا 64 سلولی , در این مراحل به جنین مورولا اطلاق می شود. مورولا توده کروی شکل حاصل از 12 تا چند صد سلول است.
بلاستولاسیون
مورولا توده توپر است ولی تسهیم ادامه یافته تا بلاستومر به سمت پیرامون خود کشیده شوند و حفره داخلی پر شده از مایع ایجاد شود. جنین یک توده از سلول هایی است که طویل نیستند ولی تشکیل یک کره را داده اند. در این مرحله جنین بلاستولا خوانده می شود و حفره پر از مایع بلاستوسل خوانده می شود. این تغییرات در حدود 10ساعت پس از لقاح به طول می انجامد. زمانی که بلاستولا برای نخست شکل می گیرد حاوی تقریبا 1000 سلول است.
گاسترولاسیون (لایه زایی)
چند ساعت کوتاه پس از بلاستولا فرایند های گاسترولاسیون (لایه زایی) آغاز می شود. در یک نقطه قطعی از سطح بلاستولا, سلولها شروع به حرکت به سمت مرکز بلاستوسل می کند. در آغاز این شکل گیری منجر به خمیدگی لوله می شود. اگر شما بادکنکی را به آرامی از آب یا هوا پر کنید و با انگشتانتان به داخل بادکنک فشار بیاوریدشما بطور تقریبی گاسرولاسیون را انجام داده اید. سطح باز شدن جنین منفذ بلاستوپور خوانده می شود و در دوتروستومیان ها (دو دهانه ای ها) نظیر توتیا و ستاره دریایی منفذ بلاستوپور یه مخرج آینده موجود تبدیل می شود. لوله خمیده آرکانترون نامیده می شود ودر نهایت به تمام جنین رسیده و در جهت دیگری از سطح جنین باز می شود و اساس شکل گیری لوله گوارشی جانور خواهد شد. از زمانی که منفذ بلاستوپور و ارکانترون شکل گیرد, جنین گاسترولا خوانده می شود.
گاسترولاسیون در جنین میکرولسیتال
در تصاویر ارایه شده، جنین در مرحله گاسترولای اولیه تا مرحله لارو" بی پی ناریا"دیده می شود.روند گاسترولاسیون مانند اندازه و توزیع زرده و تسهیم در بین گونه های مختلف در جزییات متفاوت است. در این مرحله برخی از سلول های سطح جنین به داخل حرکت کرده و لایه های سلولی جدید شکل می گیرد. این حرکات با تمایز سلول های مهاجر بداخل لایه های منفرد بافتی توام است.(ژن های خاصی فعالیت های تمایزی را باعث می شوند.)
در طی لایه زایی یک بلاستولا با خفره بلاستوسل نسبتا بزرگ شکل می گیردکه فضای کافی برای حرکت برخی از سلول های سطحی بداخل جهت تشکیل لایه های جدید را دارد.مشاهدات اولیه ستاره دریای جنین در قطب گیاهی مسطح می شود .(مهاجرت اولیه و تمایز سلول ها) که بداخل بلاستوسل قرورفتگی می یابند. و لایه سلولی داخلی شکل گرفته (هایپوبلاست یا مزودرم )و لایه های مزودرم و اندودرم شکل می گیرند.سلول های سطح خارجی گاسترولا ، اپی بلاست یا اکتودرم می شوند. و حفره بلاستوسل ، گاستروسل یاارکانترون (لوله گوارشی اولیه ) نامیده می شود. که در قطب گیاهی بعنوان بلاستوپور باز می شود.
انتهای داخلی هایپوبلاست (مزودرم) کیسه های مزودرمال را شکل می دهد. این کیسه ها Entrocelic نامیده می شوند. و اعمال انها فعالیت سلومی خوانده می شود. به موازات رشد جنین طی گاسترولاسیون ، سلول هایی از هایپوبلاست جوانه زده و برای تشکیل سلول های مزانشیمی به داخل مهاجرت میکنند.
تمایز
در مرحله گاسترولا , اندازه قطعی از تمایز نمو می یابد. سلول های سطح بیرونی گاسترولا اکتو درم را تشکیل می دهد و در آنها اپیدرم و سیستم عصبی جانور بالغ تمایز می یابد. این دولایه بوسیله لایه سوم ، مزودرم به همدیگر متصل می شوند.
در توتیا و ستاره دریایی رشد ،تمایز و شکل گیری در گاسترولا ادامه می یابد. مرحله لارو آزاد که بطور کامل قادر به حرکت و تغذیه از محیط اطراف خود است، شکل میگیرد. در مرحله بعد یک دگردیسی رخ داده و لارو به ستاره دریایی بالغ یا توتیای بالغ تبدیل می شود
تصاویری از تسهیم و لایه زایی ستاره دریایی
این صفحه یکسری از تصاویری که از تخم میکرو لیستال از سلول لقاح نیافته با مرحله بلاستولادر جنین ستاره دریایی را نشان می دهد. تخم های میکرو لیستال دارای اندازه خیلی کوچکی از زرده ذخیره شده اند. در نتیجه وجود اندازه کوچک زرده ,این حیوانات یکی از دو روش نمو را نشان میدهد.
در اولین روش لارو در مرحله جنینی از مواد تغذیه ای محیط تغذیه می شود. (در جنین های آورده شده زیر) در روش دوم که نمو داخلی است. نیاز های اساسی به وسیله مادر برآورده می شود(چیزی که ما در پستانداران مشاهده می کنیم.)
Later
تخم لقاح نیافته که از طریق اندازه و هسته مشخص (پیکان بزرگ) با هستک مشخص (پیکان کوچک) و فقدان غشای لقاحی قابل تشخیص است. تصویر زیگوت یا تخم لقاح که بوسیله غشای لقاحی (پیکانها) احاطه کرده سلول و فضای دور ویتیلینی که با مایع پر شده است،مشخص است.
Later
مرحله دو سلولی که زیگوت اولین تسهیم را کامل کرده است و تسهیم اول کامل و مساوی است.(نظیر تخمک کامل که به دو سلول تقسیم می شود). تسهیم از حول نباتی-جانوری عبور کرده است و دو بلاستومر مشابه تولید می کند. مرحله چهار سلولی که تسهیم دوم معمولا از سراسر محور گیاهی-نباتی عبور می کند. ولی بر تسهیم اول عمود است. در پایان چهار بلاستومر کامل با اندازه مساوی ایجاد می شود.
Later
مرحله هشت سلولی, سومین تسهیم در سطح استوایی رخ میدهد.(عمود بر دو تسهیم اول و محور نباتی-جانوری). چهار بلاستومر فوقانی (قطب جانوری) در اندازه کوچکتر از چهار بلاستومر تحتانی (قطب گیاهی) هستند. مرحله 32 سلولی: پس از مرحله 16 سلولی تقسیم بطور پیچیده دنبال می گردد. افزایش تعداد سلولها و تقسیمات بلاستومرها همزمان انجام می شود. تقسیم ادامه یافته و یک توده سلولی که خودش را به بلاستولا سازماندهی می کند, شکل می گیرد.
Later
بلاستولای اولیه: تسهیم ادامه پیدا می کند. در مرکز, تماس سلول ها با یکدیگر از بین رفته, و حفره مرکز پوشیده از مایع (بلاستوسل) شکل می گیرد. این بلاستوسل توسط یک لایه از سلول احاطه می شود. کره توخالی شکل گرفته به عنوان بلاستولا شناخته می شود. بلاستولای تاخیری: مشابه بلاستولای اولیه توسط یک لایه منفرد از سلول های احاطه کننده حفره مرکزی قابل تشخیص است. بلاستومرها کوچک و بطور مشخص واضح نیستند.بلاستومرهای قطب گیاهی vp ضخیم تر از قطب جانوری APظاهر می شود.
Later
گاسترولای اولیه
animal pole (AP)
vegetal pole (VP)
Later
گاسترولای میانی گاسترولای تاخیری
C
O
L
O
U
R
I
Z
E
!!
لارو بی پی ناریا
1. Blastocoel
2. Enterocoelic pouch
3. Mouth
4. Archenteron
5. Blastopore (anus) • blue=Ectoderm
• red=Mesoderm
• yellow=Endoderm
.
۱۳۸۹ آبان ۱۱, سهشنبه
gastrulation and Cleavage,in sea star.directed by:nasrin hashemi. advisor: dr minoo mahmoodi.

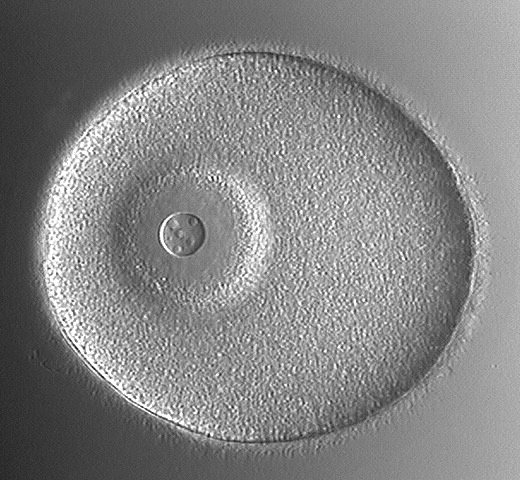


Cleavage of Microlecithal Eggs
This page shows a series of photographs which follows a microlecithal (oligolecithal) egg from the unfertilized single cell through to the late blastula. Microlecithal eggs have a very small amount of stored yolk, distributed evenly throughout the cytoplasm. As a result of this small amount of yolk, these animals show one of two different methods of development. The first is to have a feeding larval stage (as this embryo will have) to meet the nutritional requirements of the embryo, with the second being to have internal development such that the nutritional requirements of the embryo are met by the mother (what we will see later with mammals). These particular slides are whole mounts of starfish embryos.
Later
This picture is of the unfertilized egg. It can be differentiated from the zygote by the presence of a large, conspicuous nucleus (large arrow) with obvious nucleolus (smaller arrow) and by the lack of a fertilization membrane. This shows the zygote (fertilized cell). It is recognized by the presence of the fertilization membrane (arrows) surrounding it and the peripheral, fluid-filled perivitelline space.
Later
This is the two cell stage. By this stage, the zygote has completed its first cleavage, which is both equal and holoblastic (i.e. the entire ovum is divided into cells). The division (cleavage) has passed through the animal-vegetal axis, producing two similar blastomeres. This is the four cell stage. This second cleavage also passes through the animal-vegetal axis, but perpendicular to the first cleavage (described previously). Four equal-sized blastomeres are the end result.
Later
The eight cell stage. Here, the third cleavage has occurred in the equatorial plane (perpendicular to the first two cleavages and the animal-vegetal axis). Note that the upper four blastomeres (the animal pole) are slightly smaller than the lower four blastomeres (the vegetal pole). NOTE: The sixteen cell stage is not shown - its division is once again vertical, along the animal-vegetal axis. The thirty-two cell stage is seen here. After the 16 cell stage, the cleavages become more difficult to follow, due to the increasing number of cells and to the division of blastomeres becoming asynchronous. Cleavage continues, forming a mass of cells which organizes itself into the blastula. The lighter area in the centre of the embryo is the beginning of the blastocoel.
Later
This shows the early blastula. With continuing cleavage, the cells in centre begin to lose contact with one another, and a central fluid-filled cavity (the blastocoel) forms. This blastocoel is surrounded by a single layer of cells, forming the hollow sphere know as the blastula. This shows the late blastula. Like the early blastula, it is characterized by a single layer of cells surrounding the central hollow area - the blastocoel (B). The blastomeres are seen to be smaller and are individually not as obvious. The blastomeres at the vegetal pole (VP) are taller than those at the animal pole (AP), making the vegetal pole appear slightly thicker.
Gastrulation in the Microlecithal Embryo
On this page, a series of photographs can be seen which follow a microlecithal (oligolecithal) embryo from the early gastrula stage through gastrulation to the early bipinnaria larval stage.
The process of gastrulation is affected, as was cleavage, by the amount and the distribution of yolk. As a result, gastrulation differs between different species in details, but the principle is the same in all these species. During gastrulation, some of the cells from the surface of the embryo move to the interior and replicate, thereby forming new layers of cells. These movements are coupled with the differentiation of the migrating cells (caused by the differential activity of certain genes) into histologically unique layers.
During microlecithal gastrulation, a blastula with a relatively large blastocoel is usually formed. As a result, there is plenty of space to move some of the surface cells to the inside to form new layers. This is first seen as the starfish embryo becomes flattened at the vegetal pole (the initial migration and differentiation of cells), which will then invaginate within the blastocoel. When this occurs, the internal layer of cells formed (called the hypoblast or mesendoderm) are the cells which gives rise to the endoderm and mesoderm germ layers. The cells who remain on the outer surface of the gastrula become the epiblast (ectoderm). The cavity created by the invagination is called the gastrocoel or archenteron (primitive gut), whose opening at the vegetal pole is called the blastopore. The anterior end of the hypoblast (mesendoderm) develops out-pocketings (evaginations) on each side to form the mesoderm (mesodermal pouches). These pouches are also called enterocoelic pouches and the cavities within them are called the coelomic cavities. As the embryo grows during gastrulation, cells also bud off the hypoblast and migrate into the blastocoel to form mesenchymal cells.
Later
This is the early starfish gastrula. The vegetal pole (VP) is showing the first signs of invagination. AP=animal pole, VP=vegetal pole. This shows the early gastrula. The beginning of the invagination which will form the gastrocoel (a.k.a. archenteron) is obvious now, as it invaginates anteriorly (toward the animal pole (AP)) from the vegetal pole (VP), where the blastopore is formed.
Later
Here we see the mid gastrula. The gastrocoel continues to develop as the embryo grows. AP=animal pole, VP=vegetal pole. This is the late gastrula. This picture is a dorsal view of the embryo, with anterior being toward the animal pole (AP) and posterior toward the vegetal pole (VP). Note the beginning of the formation of enterocoelic pouches on the right and left side of the anterior end of the gastrocoel.
C
O
L
O
U
R
I
Z
E
!!
This is the early bipinnaria larva seen in lateral view (from the side). The dorsal aspect of the embryo is facing right and the ventral aspect is on the left side of this picture. The blastopore is at the posterior end (bottom) and is now developing into the anus, while the anterior end of the gastrocoel has bent toward the ventral aspect of the embryo, where a mouth has formed. Here we see the same picture as on the right, but now labelled and in glorious technicolour!
Blastocoel
Enterocoelic pouch
Mouth
Archenteron
Blastopore (anus) blue=Ectoderm
red=Mesoderm
yellow=Endoderm
Cleavage and Morphogenesis
Development of Sea Star Embryos (Rust, fig. 110)
Egg and Zygote
The unfertilized egg (1a) has a large and conspicuous nucleus. Compare the amount of yolk and cytoplasm to the size of the nucleus. The nucleus of the zygote (1b) is less conspicuous. The fertilization membrane is not present in this preserved material
.
cell stage
Successive cell divisions divide the zygote into smaller and more numerous cells. Compare the size of the 2,4 and 8 cell stages with the size of the original zygote. Note the cleavage furrows formed by the pinching in of the cell membranes during division. From the arrangement of the cells determine the plane of cleavage that produced the cells.
.
Early blastula
Cleavage continues producing many additional cells which arrange themselves around a fluid filled cavity (blastocoel). The blastula (7) begins to take on the appearance of a ball.
. Blastula
This ball-like stage is called the blastula (5). Compare the size of the blastula to the original zygote. Note the blastocoel (6).
late gastrula
During the late blastula stage a few cells on one side of the developing embryo begin to push into the blastocoel (8). When this process (gastrulation) becomes visible as an indentation, the developing embryo is termed a gastrula (9). The point of indentation is termed the blastopore (10). As the invagination process continues (11),with more cells pushing into the interior, a new cell layer is formed and a new cavity (12) (archenteron) appears within this new layer of cells. This cavity opens through the blastopore (14) to the outside of the embryo. Later a third layer of cells develops between the original two layers. These three cell layers (ectoderm, endoderm and mesoderm) provide the "bricks and mortar" for the differentiation of the embryo's organ systems.
. Larval
The developmental process to the end of the gastrula (13) stage takes approximately two days. At this time the gastrula transforms itself into a free swimming (via cilia bands) (18) and feeding larva (15). This change includes the formation of a digestive system with two openings (mouth (17) and anus (16)). The larva grows as it feeds on algae and diatoms. After several months the larva undergoes a process called metamorphosis that transforms it into a tiny sea star.
Sea stars belong to the Phylum Echinodermata, and display the typical deuterostome pattern of early development. In sea stars, fertilization and development take place externally and the eggs carry little yolk, so it is easy to observe early development. Obtain a slide of developing sea star embryos and
identify the following developmental stages.
_Cleavage
The earliest stages are simply named for the number of cells they contain: the 2-cell stage, the 4-cell stage, the 8-cell stage, and so on. These stages result from the first cleavage, the second cleavage, the third cleavage, etc. (Individual cells that form during these cleavages are called blastomeres (Gr: blastus = a bud, mere = part).) Once the cells become too numerous to be counted easily, which usually happens around the 32- or 64-cell stage, this terminology is abandoned and the embryo is said to be in the morula (Latin: little mulberry) stage. The morula is a spherical mass of from a few dozen to several hundred cells.
Blastulation
The morula is solid, but as cleavage continues the blastomeres move toward the periphery of the mass, leaving a fluid-filled cavity in the interior. Soon the embryo no longer resembles a simple mass of cells, but instead takes on the form of a sphere. In this stage the embryo is called a blastula (little bud), and the fluid-filled cavity is called a blastocoel (Gr: coelo = hollow). This change takes place about ten hours after fertilization. When the blastula first forms it contains about 1000 cells.
Gastrulation
A few hours after the formation of the blastula the process of gastrulation begins. At a certain point on the surface of the blastula cells begin to move toward the center of the blastocoel, so that an opening is formed leading into a blind or dead-end tube. If you were to take a rubber balloon loosely filled with water or air and poke your finger into it without breaking the balloon you would be at least roughly approximating the process of gastrulation. The opening on the surface of the embryo is called the blastopore, and in deuterostomes such as sea urchins and sea stars this will become the anus of the mature organism. The blind or dead-end tube is called the archenteron (Gr: archi = first, enterum = intestine). This will eventually reach all the way through and open out the other side, and thus will form the basis of the animal's digestive tract. Once a blastopore and an archenteron have formed, the embryo is called a gastrula (Gr: gastro = stomach).
Differentiation
At the gastrula stage of development a certain amount of differentiation may be said to have taken place. The cells on the outside of the gastrula constitute the ectoderm (Gr: ecto = outer, derma = skin), and they will develop into the epidermis and the nervous system of the mature animal. The cells of the archenteron constitute the endoderm (Gr: endo = inside), and these will become the lining of the digestive tract and the digestive glands of the fully developed organism. These two primary germ layers will be joined by a third one - the mesoderm.
In sea urchins and sea stars growth, differentiation and morphogenesis continue beyond the gastrula stage until a free-living larval stage, fully capable of moving about and feeding itself, is formed. At a later stage a metamorphosis will occur and the larva will assume the form of the adult starfish or sea urchin.
.
Highly Derived Coelomic and Water-Vascular Morphogenesis in a Starfish with Pelagic Direct Development
D. A. Janies and L. R. McEdward
University of Florida, Department of Zoology, Gainesville, Florida 32611
The coelomic development of the starfish Pteraster tesselatus (order Velatida, family Pterasteridae) is fundamentally different from that reported for all other asteroids. Coeloms arise from seven separate enterocoels that evaginate from different regions of the archenteron. The water-vascular coelomic system develops from the first five enterocoels (homologous to hydrocoel lobes) which extend radially, in a transverse orientation, from the central region of the archenteron. All other coelomic compartments derive from two enterocoels that evaginate later in development from posterior regions of the archenteron. This mode of coelom formation in P. tesselatus leads directly to the adult organization. We hypothesize that this altered pattern of coelomogenesis evolved from the pattern that occurs in the larvae of other spinulosacean asteroids, by a rotation in the site of origin of the anterior enterocoels relative to the archenteron. The altered pattern of coelomogenesis accounts for most of the unusual features of development in P. tesselatus: parallel embryonic and adult axes of symmetry, transverse orientation of the juvenile disk, absence of bilateral symmetry, absence of purely larval structures, and the lack of a metamorphosis. We conclude, contrary to previous interpretations, that P. tesselatus does not have a larval stage and thus represents the only described case of truly direct development in the asteroids.
Ultrastructural study of the hyaline layer of the starfish embryo, Pisaster ochraceus.
Campbell SS, Crawford BJ.
Department of Anatomy, University of British Columbia, Vancouver, Canada.
The hyaline layer (HL) is a multilayered extracellular matrix (ECM) that coats the external surfaces of sea urchin and starfish embryos. It is thought to protect and lubricate the embryo, stabilize the blastomeres during morphogenesis, and regulate nutrient intake. Ultrastructural studies of chemically fixed embryos have shown the HL to consist of two to four sublayers. However, since chemical fixatives may cause collapse and alter the positions and antigenicity of the extracellular components, fixation methods that exclude chemicals may reveal a picture of the HL closer to what is present in vivo. Freeze substitution, a fixation method whereby tissues are rapidly frozen and dehydrated at low temperatures, has proved useful for fixing material rich in ECM. In this study, embryos of the starfish Pisaster ochraceus were fixed for microscopy using freeze substitution and three chemical methods in order to determine, as accurately as possible, the structure of the HL. Embryos appear to be best preserved by freeze substitution and demonstrate a HL consisting of at least six distinct sublayers. Based on staining with anionic dyes, most sublayers appear to contain glycosaminoglycans. Freeze substituted embryos, which were also stained with monoclonal antibodies raised against their ECM, revealed that some molecules are common to all six sublayers, whereas other molecules may be restricted to specific sublayers. This suggests that each sublayer could have a different function. Additional evidence suggests that microvillus associated bodies, present in other marine invertebrate embryos, may anchor the asteroid HL to the cell surface microvilli.
شناسایی مراحل نمو در ستاره دریایی
تسهیم
اولین مرحله ساده که تعدادی از سلولها حاوی اند. مراحل 2 سلولی, 4سلولی, 8 سلولی و الی آخر. این مراحل از اولین , دومین, سومین وتا تسهیمات پایانی حاصل می شوند. سلولهای حاصل از هر مرحله به نام بلاستومر خوانده می شوند. سلولها به سادگی بر تعدادشان افزوده شده تا مراحل 32 سلولی یا 64 سلولی , در این مراحل به جنین مورولا اطلاق می شود. مورولا توده کروی شکل حاصل از 12 تا چند صد سلول است.
بلاستولاسیون
مورولا توده توپر است ولی تسهیم ادامه یافته تا بلاستومر به سمت پیرامون خود کشیده شوند و حفره داخلی پر شده از مایع ایجاد شود. جنین یک توده از سلول هایی است که طویل نیستند ولی تشکیل یک کره را داده اند. در این مرحله جنین بلاستولا خوانده می شود و حفره پر از مایع بلاستوسل خوانده می شود. این تغییرات در حدود 10ساعت پس از لقاح به طول می انجامد. زمانی که بلاستولا برای نخست شکل می گیرد حاوی تقریبا 1000 سلول است.
گاسترولاسیون (لایه زایی)
چند ساعت کوتاه پس از بلاستولا فرایند های گاسترولاسیون (لایه زایی) آغاز می شود. در یک نقطه قطعی از سطح بلاستولا, سلولها شروع به حرکت به سمت مرکز بلاستوسل می کند. در آغاز این شکل گیری منجر به خمیدگی لوله می شود. اگر شما بادکنکی را به آرامی از آب یا هوا پر کنید و با انگشتانتان به داخل بادکنک فشار بیاوریدشما بطور تقریبی گاسرولاسیون را انجام داده اید. سطح باز شدن جنین منفذ بلاستوپور خوانده می شود و در دوتروستومیان ها (دو دهانه ای ها) نظیر توتیا و ستاره دریایی منفذ بلاستوپور یه مخرج آینده موجود تبدیل می شود. لوله خمیده آرکانترون نامیده می شود ودر نهایت به تمام جنین رسیده و در جهت دیگری از سطح جنین باز می شود و اساس شکل گیری لوله گوارشی جانور خواهد شد. از زمانی که منفذ بلاستوپور و ارکانترون شکل گیرد, جنین گاسترولا خوانده می شود.
گاسترولاسیون در جنین میکرولسیتال
در تصاویر ارایه شده، جنین در مرحله گاسترولای اولیه تا مرحله لارو" بی پی ناریا"دیده می شود.روند گاسترولاسیون مانند اندازه و توزیع زرده و تسهیم در بین گونه های مختلف در جزییات متفاوت است. در این مرحله برخی از سلول های سطح جنین به داخل حرکت کرده و لایه های سلولی جدید شکل می گیرد. این حرکات با تمایز سلول های مهاجر بداخل لایه های منفرد بافتی توام است.(ژن های خاصی فعالیت های تمایزی را باعث می شوند.)
در طی لایه زایی یک بلاستولا با خفره بلاستوسل نسبتا بزرگ شکل می گیردکه فضای کافی برای حرکت برخی از سلول های سطحی بداخل جهت تشکیل لایه های جدید را دارد.مشاهدات اولیه ستاره دریای جنین در قطب گیاهی مسطح می شود .(مهاجرت اولیه و تمایز سلول ها) که بداخل بلاستوسل قرورفتگی می یابند. و لایه سلولی داخلی شکل گرفته (هایپوبلاست یا مزودرم )و لایه های مزودرم و اندودرم شکل می گیرند.سلول های سطح خارجی گاسترولا ، اپی بلاست یا اکتودرم می شوند. و حفره بلاستوسل ، گاستروسل یاارکانترون (لوله گوارشی اولیه ) نامیده می شود. که در قطب گیاهی بعنوان بلاستوپور باز می شود.
انتهای داخلی هایپوبلاست (مزودرم) کیسه های مزودرمال را شکل می دهد. این کیسه ها Entrocelic نامیده می شوند. و اعمال انها فعالیت سلومی خوانده می شود. به موازات رشد جنین طی گاسترولاسیون ، سلول هایی از هایپوبلاست جوانه زده و برای تشکیل سلول های مزانشیمی به داخل مهاجرت میکنند.
تمایز
در مرحله گاسترولا , اندازه قطعی از تمایز نمو می یابد. سلول های سطح بیرونی گاسترولا اکتو درم را تشکیل می دهد و در آنها اپیدرم و سیستم عصبی جانور بالغ تمایز می یابد. این دولایه بوسیله لایه سوم ، مزودرم به همدیگر متصل می شوند.
در توتیا و ستاره دریایی رشد ،تمایز و شکل گیری در گاسترولا ادامه می یابد. مرحله لارو آزاد که بطور کامل قادر به حرکت و تغذیه از محیط اطراف خود است، شکل میگیرد. در مرحله بعد یک دگردیسی رخ داده و لارو به ستاره دریایی بالغ یا توتیای بالغ تبدیل می شود
تصاویری از تسهیم و لایه زایی ستاره دریایی
این صفحه یکسری از تصاویری که از تخم میکرو لیستال از سلول لقاح نیافته با مرحله بلاستولادر جنین ستاره دریایی را نشان می دهد. تخم های میکرو لیستال دارای اندازه خیلی کوچکی از زرده ذخیره شده اند. در نتیجه وجود اندازه کوچک زرده ,این حیوانات یکی از دو روش نمو را نشان میدهد.
در اولین روش لارو در مرحله جنینی از مواد تغذیه ای محیط تغذیه می شود. (در جنین های آورده شده زیر) در روش دوم که نمو داخلی است. نیاز های اساسی به وسیله مادر برآورده می شود(چیزی که ما در پستانداران مشاهده می کنیم.)
Later
تخم لقاح نیافته که از طریق اندازه و هسته مشخص (پیکان بزرگ) با هستک مشخص (پیکان کوچک) و فقدان غشای لقاحی قابل تشخیص است. تصویر زیگوت یا تخم لقاح که بوسیله غشای لقاحی (پیکانها) احاطه کرده سلول و فضای دور ویتیلینی که با مایع پر شده است،مشخص است.
Later
مرحله دو سلولی که زیگوت اولین تسهیم را کامل کرده است و تسهیم اول کامل و مساوی است.(نظیر تخمک کامل که به دو سلول تقسیم می شود). تسهیم از حول نباتی-جانوری عبور کرده است و دو بلاستومر مشابه تولید می کند. مرحله چهار سلولی که تسهیم دوم معمولا از سراسر محور گیاهی-نباتی عبور می کند. ولی بر تسهیم اول عمود است. در پایان چهار بلاستومر کامل با اندازه مساوی ایجاد می شود.
Later
مرحله هشت سلولی, سومین تسهیم در سطح استوایی رخ میدهد.(عمود بر دو تسهیم اول و محور نباتی-جانوری). چهار بلاستومر فوقانی (قطب جانوری) در اندازه کوچکتر از چهار بلاستومر تحتانی (قطب گیاهی) هستند. مرحله 32 سلولی: پس از مرحله 16 سلولی تقسیم بطور پیچیده دنبال می گردد. افزایش تعداد سلولها و تقسیمات بلاستومرها همزمان انجام می شود. تقسیم ادامه یافته و یک توده سلولی که خودش را به بلاستولا سازماندهی می کند, شکل می گیرد.
Later
بلاستولای اولیه: تسهیم ادامه پیدا می کند. در مرکز, تماس سلول ها با یکدیگر از بین رفته, و حفره مرکز پوشیده از مایع (بلاستوسل) شکل می گیرد. این بلاستوسل توسط یک لایه از سلول احاطه می شود. کره توخالی شکل گرفته به عنوان بلاستولا شناخته می شود. بلاستولای تاخیری: مشابه بلاستولای اولیه توسط یک لایه منفرد از سلول های احاطه کننده حفره مرکزی قابل تشخیص است. بلاستومرها کوچک و بطور مشخص واضح نیستند.بلاستومرهای قطب گیاهی vp ضخیم تر از قطب جانوری APظاهر می شود.
Later
گاسترولای اولیه
animal pole (AP)
vegetal pole (VP)
Later
گاسترولای میانی گاسترولای تاخیری
C
O
L
O
U
R
I
Z
E
!!
لارو بی پی ناریا
1. Blastocoel
2. Enterocoelic pouch
3. Mouth
4. Archenteron
5. Blastopore (anus) • blue=Ectoderm
• red=Mesoderm
• yellow=Endoderm
.
This page shows a series of photographs which follows a microlecithal (oligolecithal) egg from the unfertilized single cell through to the late blastula. Microlecithal eggs have a very small amount of stored yolk, distributed evenly throughout the cytoplasm. As a result of this small amount of yolk, these animals show one of two different methods of development. The first is to have a feeding larval stage (as this embryo will have) to meet the nutritional requirements of the embryo, with the second being to have internal development such that the nutritional requirements of the embryo are met by the mother (what we will see later with mammals). These particular slides are whole mounts of starfish embryos.
Later
This picture is of the unfertilized egg. It can be differentiated from the zygote by the presence of a large, conspicuous nucleus (large arrow) with obvious nucleolus (smaller arrow) and by the lack of a fertilization membrane. This shows the zygote (fertilized cell). It is recognized by the presence of the fertilization membrane (arrows) surrounding it and the peripheral, fluid-filled perivitelline space.
Later
This is the two cell stage. By this stage, the zygote has completed its first cleavage, which is both equal and holoblastic (i.e. the entire ovum is divided into cells). The division (cleavage) has passed through the animal-vegetal axis, producing two similar blastomeres. This is the four cell stage. This second cleavage also passes through the animal-vegetal axis, but perpendicular to the first cleavage (described previously). Four equal-sized blastomeres are the end result.
Later
The eight cell stage. Here, the third cleavage has occurred in the equatorial plane (perpendicular to the first two cleavages and the animal-vegetal axis). Note that the upper four blastomeres (the animal pole) are slightly smaller than the lower four blastomeres (the vegetal pole). NOTE: The sixteen cell stage is not shown - its division is once again vertical, along the animal-vegetal axis. The thirty-two cell stage is seen here. After the 16 cell stage, the cleavages become more difficult to follow, due to the increasing number of cells and to the division of blastomeres becoming asynchronous. Cleavage continues, forming a mass of cells which organizes itself into the blastula. The lighter area in the centre of the embryo is the beginning of the blastocoel.
Later
This shows the early blastula. With continuing cleavage, the cells in centre begin to lose contact with one another, and a central fluid-filled cavity (the blastocoel) forms. This blastocoel is surrounded by a single layer of cells, forming the hollow sphere know as the blastula. This shows the late blastula. Like the early blastula, it is characterized by a single layer of cells surrounding the central hollow area - the blastocoel (B). The blastomeres are seen to be smaller and are individually not as obvious. The blastomeres at the vegetal pole (VP) are taller than those at the animal pole (AP), making the vegetal pole appear slightly thicker.
Gastrulation in the Microlecithal Embryo
On this page, a series of photographs can be seen which follow a microlecithal (oligolecithal) embryo from the early gastrula stage through gastrulation to the early bipinnaria larval stage.
The process of gastrulation is affected, as was cleavage, by the amount and the distribution of yolk. As a result, gastrulation differs between different species in details, but the principle is the same in all these species. During gastrulation, some of the cells from the surface of the embryo move to the interior and replicate, thereby forming new layers of cells. These movements are coupled with the differentiation of the migrating cells (caused by the differential activity of certain genes) into histologically unique layers.
During microlecithal gastrulation, a blastula with a relatively large blastocoel is usually formed. As a result, there is plenty of space to move some of the surface cells to the inside to form new layers. This is first seen as the starfish embryo becomes flattened at the vegetal pole (the initial migration and differentiation of cells), which will then invaginate within the blastocoel. When this occurs, the internal layer of cells formed (called the hypoblast or mesendoderm) are the cells which gives rise to the endoderm and mesoderm germ layers. The cells who remain on the outer surface of the gastrula become the epiblast (ectoderm). The cavity created by the invagination is called the gastrocoel or archenteron (primitive gut), whose opening at the vegetal pole is called the blastopore. The anterior end of the hypoblast (mesendoderm) develops out-pocketings (evaginations) on each side to form the mesoderm (mesodermal pouches). These pouches are also called enterocoelic pouches and the cavities within them are called the coelomic cavities. As the embryo grows during gastrulation, cells also bud off the hypoblast and migrate into the blastocoel to form mesenchymal cells.
Later
This is the early starfish gastrula. The vegetal pole (VP) is showing the first signs of invagination. AP=animal pole, VP=vegetal pole. This shows the early gastrula. The beginning of the invagination which will form the gastrocoel (a.k.a. archenteron) is obvious now, as it invaginates anteriorly (toward the animal pole (AP)) from the vegetal pole (VP), where the blastopore is formed.
Later
Here we see the mid gastrula. The gastrocoel continues to develop as the embryo grows. AP=animal pole, VP=vegetal pole. This is the late gastrula. This picture is a dorsal view of the embryo, with anterior being toward the animal pole (AP) and posterior toward the vegetal pole (VP). Note the beginning of the formation of enterocoelic pouches on the right and left side of the anterior end of the gastrocoel.
C
O
L
O
U
R
I
Z
E
!!
This is the early bipinnaria larva seen in lateral view (from the side). The dorsal aspect of the embryo is facing right and the ventral aspect is on the left side of this picture. The blastopore is at the posterior end (bottom) and is now developing into the anus, while the anterior end of the gastrocoel has bent toward the ventral aspect of the embryo, where a mouth has formed. Here we see the same picture as on the right, but now labelled and in glorious technicolour!
Blastocoel
Enterocoelic pouch
Mouth
Archenteron
Blastopore (anus) blue=Ectoderm
red=Mesoderm
yellow=Endoderm
Cleavage and Morphogenesis
Development of Sea Star Embryos (Rust, fig. 110)
Egg and Zygote
The unfertilized egg (1a) has a large and conspicuous nucleus. Compare the amount of yolk and cytoplasm to the size of the nucleus. The nucleus of the zygote (1b) is less conspicuous. The fertilization membrane is not present in this preserved material
.
cell stage
Successive cell divisions divide the zygote into smaller and more numerous cells. Compare the size of the 2,4 and 8 cell stages with the size of the original zygote. Note the cleavage furrows formed by the pinching in of the cell membranes during division. From the arrangement of the cells determine the plane of cleavage that produced the cells.
.
Early blastula
Cleavage continues producing many additional cells which arrange themselves around a fluid filled cavity (blastocoel). The blastula (7) begins to take on the appearance of a ball.
. Blastula
This ball-like stage is called the blastula (5). Compare the size of the blastula to the original zygote. Note the blastocoel (6).
late gastrula
During the late blastula stage a few cells on one side of the developing embryo begin to push into the blastocoel (8). When this process (gastrulation) becomes visible as an indentation, the developing embryo is termed a gastrula (9). The point of indentation is termed the blastopore (10). As the invagination process continues (11),with more cells pushing into the interior, a new cell layer is formed and a new cavity (12) (archenteron) appears within this new layer of cells. This cavity opens through the blastopore (14) to the outside of the embryo. Later a third layer of cells develops between the original two layers. These three cell layers (ectoderm, endoderm and mesoderm) provide the "bricks and mortar" for the differentiation of the embryo's organ systems.
. Larval
The developmental process to the end of the gastrula (13) stage takes approximately two days. At this time the gastrula transforms itself into a free swimming (via cilia bands) (18) and feeding larva (15). This change includes the formation of a digestive system with two openings (mouth (17) and anus (16)). The larva grows as it feeds on algae and diatoms. After several months the larva undergoes a process called metamorphosis that transforms it into a tiny sea star.
Sea stars belong to the Phylum Echinodermata, and display the typical deuterostome pattern of early development. In sea stars, fertilization and development take place externally and the eggs carry little yolk, so it is easy to observe early development. Obtain a slide of developing sea star embryos and
identify the following developmental stages.
_Cleavage
The earliest stages are simply named for the number of cells they contain: the 2-cell stage, the 4-cell stage, the 8-cell stage, and so on. These stages result from the first cleavage, the second cleavage, the third cleavage, etc. (Individual cells that form during these cleavages are called blastomeres (Gr: blastus = a bud, mere = part).) Once the cells become too numerous to be counted easily, which usually happens around the 32- or 64-cell stage, this terminology is abandoned and the embryo is said to be in the morula (Latin: little mulberry) stage. The morula is a spherical mass of from a few dozen to several hundred cells.
Blastulation
The morula is solid, but as cleavage continues the blastomeres move toward the periphery of the mass, leaving a fluid-filled cavity in the interior. Soon the embryo no longer resembles a simple mass of cells, but instead takes on the form of a sphere. In this stage the embryo is called a blastula (little bud), and the fluid-filled cavity is called a blastocoel (Gr: coelo = hollow). This change takes place about ten hours after fertilization. When the blastula first forms it contains about 1000 cells.
Gastrulation
A few hours after the formation of the blastula the process of gastrulation begins. At a certain point on the surface of the blastula cells begin to move toward the center of the blastocoel, so that an opening is formed leading into a blind or dead-end tube. If you were to take a rubber balloon loosely filled with water or air and poke your finger into it without breaking the balloon you would be at least roughly approximating the process of gastrulation. The opening on the surface of the embryo is called the blastopore, and in deuterostomes such as sea urchins and sea stars this will become the anus of the mature organism. The blind or dead-end tube is called the archenteron (Gr: archi = first, enterum = intestine). This will eventually reach all the way through and open out the other side, and thus will form the basis of the animal's digestive tract. Once a blastopore and an archenteron have formed, the embryo is called a gastrula (Gr: gastro = stomach).
Differentiation
At the gastrula stage of development a certain amount of differentiation may be said to have taken place. The cells on the outside of the gastrula constitute the ectoderm (Gr: ecto = outer, derma = skin), and they will develop into the epidermis and the nervous system of the mature animal. The cells of the archenteron constitute the endoderm (Gr: endo = inside), and these will become the lining of the digestive tract and the digestive glands of the fully developed organism. These two primary germ layers will be joined by a third one - the mesoderm.
In sea urchins and sea stars growth, differentiation and morphogenesis continue beyond the gastrula stage until a free-living larval stage, fully capable of moving about and feeding itself, is formed. At a later stage a metamorphosis will occur and the larva will assume the form of the adult starfish or sea urchin.
.
Highly Derived Coelomic and Water-Vascular Morphogenesis in a Starfish with Pelagic Direct Development
D. A. Janies and L. R. McEdward
University of Florida, Department of Zoology, Gainesville, Florida 32611
The coelomic development of the starfish Pteraster tesselatus (order Velatida, family Pterasteridae) is fundamentally different from that reported for all other asteroids. Coeloms arise from seven separate enterocoels that evaginate from different regions of the archenteron. The water-vascular coelomic system develops from the first five enterocoels (homologous to hydrocoel lobes) which extend radially, in a transverse orientation, from the central region of the archenteron. All other coelomic compartments derive from two enterocoels that evaginate later in development from posterior regions of the archenteron. This mode of coelom formation in P. tesselatus leads directly to the adult organization. We hypothesize that this altered pattern of coelomogenesis evolved from the pattern that occurs in the larvae of other spinulosacean asteroids, by a rotation in the site of origin of the anterior enterocoels relative to the archenteron. The altered pattern of coelomogenesis accounts for most of the unusual features of development in P. tesselatus: parallel embryonic and adult axes of symmetry, transverse orientation of the juvenile disk, absence of bilateral symmetry, absence of purely larval structures, and the lack of a metamorphosis. We conclude, contrary to previous interpretations, that P. tesselatus does not have a larval stage and thus represents the only described case of truly direct development in the asteroids.
Ultrastructural study of the hyaline layer of the starfish embryo, Pisaster ochraceus.
Campbell SS, Crawford BJ.
Department of Anatomy, University of British Columbia, Vancouver, Canada.
The hyaline layer (HL) is a multilayered extracellular matrix (ECM) that coats the external surfaces of sea urchin and starfish embryos. It is thought to protect and lubricate the embryo, stabilize the blastomeres during morphogenesis, and regulate nutrient intake. Ultrastructural studies of chemically fixed embryos have shown the HL to consist of two to four sublayers. However, since chemical fixatives may cause collapse and alter the positions and antigenicity of the extracellular components, fixation methods that exclude chemicals may reveal a picture of the HL closer to what is present in vivo. Freeze substitution, a fixation method whereby tissues are rapidly frozen and dehydrated at low temperatures, has proved useful for fixing material rich in ECM. In this study, embryos of the starfish Pisaster ochraceus were fixed for microscopy using freeze substitution and three chemical methods in order to determine, as accurately as possible, the structure of the HL. Embryos appear to be best preserved by freeze substitution and demonstrate a HL consisting of at least six distinct sublayers. Based on staining with anionic dyes, most sublayers appear to contain glycosaminoglycans. Freeze substituted embryos, which were also stained with monoclonal antibodies raised against their ECM, revealed that some molecules are common to all six sublayers, whereas other molecules may be restricted to specific sublayers. This suggests that each sublayer could have a different function. Additional evidence suggests that microvillus associated bodies, present in other marine invertebrate embryos, may anchor the asteroid HL to the cell surface microvilli.
شناسایی مراحل نمو در ستاره دریایی
تسهیم
اولین مرحله ساده که تعدادی از سلولها حاوی اند. مراحل 2 سلولی, 4سلولی, 8 سلولی و الی آخر. این مراحل از اولین , دومین, سومین وتا تسهیمات پایانی حاصل می شوند. سلولهای حاصل از هر مرحله به نام بلاستومر خوانده می شوند. سلولها به سادگی بر تعدادشان افزوده شده تا مراحل 32 سلولی یا 64 سلولی , در این مراحل به جنین مورولا اطلاق می شود. مورولا توده کروی شکل حاصل از 12 تا چند صد سلول است.
بلاستولاسیون
مورولا توده توپر است ولی تسهیم ادامه یافته تا بلاستومر به سمت پیرامون خود کشیده شوند و حفره داخلی پر شده از مایع ایجاد شود. جنین یک توده از سلول هایی است که طویل نیستند ولی تشکیل یک کره را داده اند. در این مرحله جنین بلاستولا خوانده می شود و حفره پر از مایع بلاستوسل خوانده می شود. این تغییرات در حدود 10ساعت پس از لقاح به طول می انجامد. زمانی که بلاستولا برای نخست شکل می گیرد حاوی تقریبا 1000 سلول است.
گاسترولاسیون (لایه زایی)
چند ساعت کوتاه پس از بلاستولا فرایند های گاسترولاسیون (لایه زایی) آغاز می شود. در یک نقطه قطعی از سطح بلاستولا, سلولها شروع به حرکت به سمت مرکز بلاستوسل می کند. در آغاز این شکل گیری منجر به خمیدگی لوله می شود. اگر شما بادکنکی را به آرامی از آب یا هوا پر کنید و با انگشتانتان به داخل بادکنک فشار بیاوریدشما بطور تقریبی گاسرولاسیون را انجام داده اید. سطح باز شدن جنین منفذ بلاستوپور خوانده می شود و در دوتروستومیان ها (دو دهانه ای ها) نظیر توتیا و ستاره دریایی منفذ بلاستوپور یه مخرج آینده موجود تبدیل می شود. لوله خمیده آرکانترون نامیده می شود ودر نهایت به تمام جنین رسیده و در جهت دیگری از سطح جنین باز می شود و اساس شکل گیری لوله گوارشی جانور خواهد شد. از زمانی که منفذ بلاستوپور و ارکانترون شکل گیرد, جنین گاسترولا خوانده می شود.
گاسترولاسیون در جنین میکرولسیتال
در تصاویر ارایه شده، جنین در مرحله گاسترولای اولیه تا مرحله لارو" بی پی ناریا"دیده می شود.روند گاسترولاسیون مانند اندازه و توزیع زرده و تسهیم در بین گونه های مختلف در جزییات متفاوت است. در این مرحله برخی از سلول های سطح جنین به داخل حرکت کرده و لایه های سلولی جدید شکل می گیرد. این حرکات با تمایز سلول های مهاجر بداخل لایه های منفرد بافتی توام است.(ژن های خاصی فعالیت های تمایزی را باعث می شوند.)
در طی لایه زایی یک بلاستولا با خفره بلاستوسل نسبتا بزرگ شکل می گیردکه فضای کافی برای حرکت برخی از سلول های سطحی بداخل جهت تشکیل لایه های جدید را دارد.مشاهدات اولیه ستاره دریای جنین در قطب گیاهی مسطح می شود .(مهاجرت اولیه و تمایز سلول ها) که بداخل بلاستوسل قرورفتگی می یابند. و لایه سلولی داخلی شکل گرفته (هایپوبلاست یا مزودرم )و لایه های مزودرم و اندودرم شکل می گیرند.سلول های سطح خارجی گاسترولا ، اپی بلاست یا اکتودرم می شوند. و حفره بلاستوسل ، گاستروسل یاارکانترون (لوله گوارشی اولیه ) نامیده می شود. که در قطب گیاهی بعنوان بلاستوپور باز می شود.
انتهای داخلی هایپوبلاست (مزودرم) کیسه های مزودرمال را شکل می دهد. این کیسه ها Entrocelic نامیده می شوند. و اعمال انها فعالیت سلومی خوانده می شود. به موازات رشد جنین طی گاسترولاسیون ، سلول هایی از هایپوبلاست جوانه زده و برای تشکیل سلول های مزانشیمی به داخل مهاجرت میکنند.
تمایز
در مرحله گاسترولا , اندازه قطعی از تمایز نمو می یابد. سلول های سطح بیرونی گاسترولا اکتو درم را تشکیل می دهد و در آنها اپیدرم و سیستم عصبی جانور بالغ تمایز می یابد. این دولایه بوسیله لایه سوم ، مزودرم به همدیگر متصل می شوند.
در توتیا و ستاره دریایی رشد ،تمایز و شکل گیری در گاسترولا ادامه می یابد. مرحله لارو آزاد که بطور کامل قادر به حرکت و تغذیه از محیط اطراف خود است، شکل میگیرد. در مرحله بعد یک دگردیسی رخ داده و لارو به ستاره دریایی بالغ یا توتیای بالغ تبدیل می شود
تصاویری از تسهیم و لایه زایی ستاره دریایی
این صفحه یکسری از تصاویری که از تخم میکرو لیستال از سلول لقاح نیافته با مرحله بلاستولادر جنین ستاره دریایی را نشان می دهد. تخم های میکرو لیستال دارای اندازه خیلی کوچکی از زرده ذخیره شده اند. در نتیجه وجود اندازه کوچک زرده ,این حیوانات یکی از دو روش نمو را نشان میدهد.
در اولین روش لارو در مرحله جنینی از مواد تغذیه ای محیط تغذیه می شود. (در جنین های آورده شده زیر) در روش دوم که نمو داخلی است. نیاز های اساسی به وسیله مادر برآورده می شود(چیزی که ما در پستانداران مشاهده می کنیم.)
Later
تخم لقاح نیافته که از طریق اندازه و هسته مشخص (پیکان بزرگ) با هستک مشخص (پیکان کوچک) و فقدان غشای لقاحی قابل تشخیص است. تصویر زیگوت یا تخم لقاح که بوسیله غشای لقاحی (پیکانها) احاطه کرده سلول و فضای دور ویتیلینی که با مایع پر شده است،مشخص است.
Later
مرحله دو سلولی که زیگوت اولین تسهیم را کامل کرده است و تسهیم اول کامل و مساوی است.(نظیر تخمک کامل که به دو سلول تقسیم می شود). تسهیم از حول نباتی-جانوری عبور کرده است و دو بلاستومر مشابه تولید می کند. مرحله چهار سلولی که تسهیم دوم معمولا از سراسر محور گیاهی-نباتی عبور می کند. ولی بر تسهیم اول عمود است. در پایان چهار بلاستومر کامل با اندازه مساوی ایجاد می شود.
Later
مرحله هشت سلولی, سومین تسهیم در سطح استوایی رخ میدهد.(عمود بر دو تسهیم اول و محور نباتی-جانوری). چهار بلاستومر فوقانی (قطب جانوری) در اندازه کوچکتر از چهار بلاستومر تحتانی (قطب گیاهی) هستند. مرحله 32 سلولی: پس از مرحله 16 سلولی تقسیم بطور پیچیده دنبال می گردد. افزایش تعداد سلولها و تقسیمات بلاستومرها همزمان انجام می شود. تقسیم ادامه یافته و یک توده سلولی که خودش را به بلاستولا سازماندهی می کند, شکل می گیرد.
Later
بلاستولای اولیه: تسهیم ادامه پیدا می کند. در مرکز, تماس سلول ها با یکدیگر از بین رفته, و حفره مرکز پوشیده از مایع (بلاستوسل) شکل می گیرد. این بلاستوسل توسط یک لایه از سلول احاطه می شود. کره توخالی شکل گرفته به عنوان بلاستولا شناخته می شود. بلاستولای تاخیری: مشابه بلاستولای اولیه توسط یک لایه منفرد از سلول های احاطه کننده حفره مرکزی قابل تشخیص است. بلاستومرها کوچک و بطور مشخص واضح نیستند.بلاستومرهای قطب گیاهی vp ضخیم تر از قطب جانوری APظاهر می شود.
Later
گاسترولای اولیه
animal pole (AP)
vegetal pole (VP)
Later
گاسترولای میانی گاسترولای تاخیری
C
O
L
O
U
R
I
Z
E
!!
لارو بی پی ناریا
1. Blastocoel
2. Enterocoelic pouch
3. Mouth
4. Archenteron
5. Blastopore (anus) • blue=Ectoderm
• red=Mesoderm
• yellow=Endoderm
.
The Evolution of the Eye!!! dr mahmodi


directed by:mohamad mehdi shafiei
Types of Eyes
The Simple Eye (3 Main Kinds):
1. The Positive Lens or Camera Eye Found in vertebrates and some invertebrates. This is the type of eye humans have. You can even poke it with your finger right now.
The Simple Eye
2. The Concave Mirror Eye
Found in the clam Pecten and a few ostracod crustaceans. This produces bright but reasonably hazy picture.
3. The Pinhole Eye
Pit or Cup eyes are found mainly in mollusks and can only resolve location of objects.
Made up of ommatidia receptors, each of which functions as a separate visual receptor.
The Apposition Eye
Ommatidia function independently.
The Superposition Eye
Ommatidia cooperate to produce a brighter, superimposed image on the retina.
The Two Kinds of Photoreceptors
All photoreceptors use a light sensitive pigment derived from vitamin A which is bound to an opsin. After being exposing the photopigment to light the opsin binds to a G-protein (common neurotransmitter). These similarities suggest a shared ancestry.
Rhabdomeric Photoreceptors (in protosomes)
Found mainly in the compund eyes of arthropods.
Ciliary Photoreceptors (in deuterosomes)
Common in vertebrates.
Rhabdomeric Photoreceptors
Increase their surface area by throwing up their apical surfaces into numerous folds (think Bart Simpson’s hair). Many morphologies for this receptor exist.
Ciliary Photoreceptor
Increases membrane surface area by modifying the cilium. The ciliary membrane is expanded and thrown into deep folds, so that the actuala receptor region of the cell looks like a stack of discs.
Evidence of Common Receptor Ancestry
Rods and Cones seem to have evolved from common ciliary photoreceptor precursors, while retinal ganglion, amacrine and horizontal cells seem to have evolved from a rhabdomeric precursor.
How Could Eyes Evolve?
Creationists often use the eye as a debate point to show that evolution is flawed, citing that the eye is too complex and perfect to have evolved. Darwin himself noted that “To suppose that the eye… could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree,” (The Origin of Species). He follows, however, with the assertion that eyes could likely evolve from light sensitive neurons. This seems to be the case.
The Evolution of the Eye
Research by Dan-E Nilsson and Susanne Pelger indicates that it is in fact easier to estimate the number of generations necessary to evolve an eye than complex organs. This is because these changes can be viewed as quantitative local modifications to a pre-existing tissue.
In order to determine the number of generations needed to evolve an eye, Nilsson simply made calculations outlining the plausible sequence of alterations leading from a light sensitive spot to a fully developed lens eye.
The Evolution of the Eye
Nilsson assumed an organism with a light sensitive patch of cells resting on a dark pigmented background and placed in a selection for spatial recognition. The first method to create a spatial recognition is either for a depression to form in the center of the patch, or for the edges of the patch to constrict and raise. This cupping would allow for the vague correlation of light to position where the exposure of an area on the patch is dependent on its angle to the light source.
The Evolution of the Eye
This cupping evolution should first favor the formation of a depression in the patch, than the constriction of an aperture via the raising and constriction of the surrounding pigment epithelium. This results in a sunken eye cup that resembles that of some mollusks.
This pinhole-like eye is not very good at resolving detail and creates a very dim image. Because of this, any change that improves clarity and illumination will be favored. The two routes of change for this would be the development of a lens, or the increase in the size of the eye. Increasing the size of the eye, however, presents physical problems and less acute vision than a lens would.
The Simple Eye (3 Main Kinds):
1. The Positive Lens or Camera Eye Found in vertebrates and some invertebrates. This is the type of eye humans have. You can even poke it with your finger right now.
The Simple Eye
2. The Concave Mirror Eye
Found in the clam Pecten and a few ostracod crustaceans. This produces bright but reasonably hazy picture.
3. The Pinhole Eye
Pit or Cup eyes are found mainly in mollusks and can only resolve location of objects.
Made up of ommatidia receptors, each of which functions as a separate visual receptor.
The Apposition Eye
Ommatidia function independently.
The Superposition Eye
Ommatidia cooperate to produce a brighter, superimposed image on the retina.
The Two Kinds of Photoreceptors
All photoreceptors use a light sensitive pigment derived from vitamin A which is bound to an opsin. After being exposing the photopigment to light the opsin binds to a G-protein (common neurotransmitter). These similarities suggest a shared ancestry.
Rhabdomeric Photoreceptors (in protosomes)
Found mainly in the compund eyes of arthropods.
Ciliary Photoreceptors (in deuterosomes)
Common in vertebrates.
Rhabdomeric Photoreceptors
Increase their surface area by throwing up their apical surfaces into numerous folds (think Bart Simpson’s hair). Many morphologies for this receptor exist.
Ciliary Photoreceptor
Increases membrane surface area by modifying the cilium. The ciliary membrane is expanded and thrown into deep folds, so that the actuala receptor region of the cell looks like a stack of discs.
Evidence of Common Receptor Ancestry
Rods and Cones seem to have evolved from common ciliary photoreceptor precursors, while retinal ganglion, amacrine and horizontal cells seem to have evolved from a rhabdomeric precursor.
How Could Eyes Evolve?
Creationists often use the eye as a debate point to show that evolution is flawed, citing that the eye is too complex and perfect to have evolved. Darwin himself noted that “To suppose that the eye… could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree,” (The Origin of Species). He follows, however, with the assertion that eyes could likely evolve from light sensitive neurons. This seems to be the case.
The Evolution of the Eye
Research by Dan-E Nilsson and Susanne Pelger indicates that it is in fact easier to estimate the number of generations necessary to evolve an eye than complex organs. This is because these changes can be viewed as quantitative local modifications to a pre-existing tissue.
In order to determine the number of generations needed to evolve an eye, Nilsson simply made calculations outlining the plausible sequence of alterations leading from a light sensitive spot to a fully developed lens eye.
The Evolution of the Eye
Nilsson assumed an organism with a light sensitive patch of cells resting on a dark pigmented background and placed in a selection for spatial recognition. The first method to create a spatial recognition is either for a depression to form in the center of the patch, or for the edges of the patch to constrict and raise. This cupping would allow for the vague correlation of light to position where the exposure of an area on the patch is dependent on its angle to the light source.
The Evolution of the Eye
This cupping evolution should first favor the formation of a depression in the patch, than the constriction of an aperture via the raising and constriction of the surrounding pigment epithelium. This results in a sunken eye cup that resembles that of some mollusks.
This pinhole-like eye is not very good at resolving detail and creates a very dim image. Because of this, any change that improves clarity and illumination will be favored. The two routes of change for this would be the development of a lens, or the increase in the size of the eye. Increasing the size of the eye, however, presents physical problems and less acute vision than a lens would.
تكامل چشم.کاری از آقای محمد مهدی شفیعی. نام استاد : دکتر محمودی
تكامل چشم
انواع چشم:
چشم معمولي سه نوع است:
1- لنز ساده يا چشم دوربين: در مهره داران و برخي از بي مهرگان يافت مي شود. اين نوع از چشم را انسان داراست. حتي شما مي توانيد همين الان با انگشتتان به آن فشار وارد كنيد. ادامه بدهيد، آرام . همينه.
2- چشم آينه اي مقعر: در حلزون دو كپه اي يا صدف خوراكي از جنس pecten و تعداد كمي سخت پوستان دو كپه اي و خرچمگ ها يافت مي شود. اين نوع چشم براي جانور توليد روشنايي و تصاوير مبهم مي كند.
3- The pinhale Eye: چشم هاي حفره دار يا سوراخ عمدتاً در نرم تنان يافت مي شود و فقطمي توانند مكان اشياء را تشخيص دهند.
چشم مركب:
كه از دريافت كننده هاي چشم هاي سادة حشرات تشكيل شده و هر كداك به عنوان يك دريافت كننده (رسپنتور) بينايي جداگانه عمل مي كنند.
چشم متضاد: چشم هاي سادة حشرات به طور مستقل عمل مي كنند. (غيروابسته)
The superposition Eye: چشم هاي سادة حشرات براي توليد تصوير بزرگتر و با جزئيات بيشتر بر روي شبكيه با هم همكاري مي كنند.
دو نوع رسپنتورهاي حساس به نور مرئي:
همة رسپنتورهاي حساس به نور از يك رنگدانة حساس به نور كه از ويتاكين A تركيب شده با opsin مشتق شده است، استفاده مي كنند. بعد از اين كه رنگدانة نوري در معرض روشنايي قرار گرفت، opsin به يك G – پروتئين (نروترانسميتر متداول) پيوند مي خورد. اين مشابهت ها يك ريشه مشترك را پيشنهاد مي كند.
گيرنده هاي نوري: Rhabdomeric (در protosomes) كه عمدتاً در چشمان بندپايان يافت مي شود.
گيرنده هاي نوري مژك دار (در deuterosomes) در مهره داران مشترك است.
Rhabdomeric photoreceptors :
مناطق سطحي خود را به وسيلة بلند كردن سطوح قله اي در چين هاي بسيار زياد، افزايش مي دهد. (نظرية موي بارت سيمپسون). ريخت شناسي هاي بسياري براي اين گيرنده موجود است.
گيرنده هاي نوري مژك دار:
مناطق سطحي غشاء را به وسيلة تعديل مژك ها افزايش مي دهد. غشاي مژك دار متسع شده و وارد چين هاي عميق مي شود. در نتيجه ناحية واقعي گيرندة سلول مانند بسته هاي ديسك مي شود.
ميله ها و مخروط ها به نظر مي رسد كه از مواد متشكله متداول گيرندة نوري مژك دار منشأ گرفته و استنتاج شده اند، در حالي كه عقدة شبكيه اي، amacrine و سلول هاي افقي به نظر مي رسد كه از يك ماده متشكله Rhobdonieric استنتاج شده است.
چشم ها چگونه تكامل يافته اند؟
متخصصان افرينش اغلب از چشم به عنوان يك نقطه بحث و مناظره استفاده مي كنند تا نشان دهند كه تكامل نقض شده است و ذكر كرده اند كه چشم خيلي پيچيده است و از نظر تكاملي عاليتر از تكامل يافته است. خود «داروين» نوشته است كه «براي فرض كردن اين كه چشم مي تواند به وسيلة گزينش هاي طبيعي تشكيل شده باشد، به نظر مي رسد، من آزادانه اعتراف مي كنم كه اين بحث نامعقولي در بالاترين حد ممكن است. او در ادامه به هر حال، با تأكيد گفت: چشم ها احتمالاً از نرون هاي حساس به روشنايي تكامل يافته اند، به نظر مي رسد كه واقعيت باشد.
تكامل چشم:
تحقيقات دَن اِ نيلسون و سوزان بِلگِر نشان مي دهد كه در حقيقت تخمين تعداد توليدات ضروري براي تكامل يك چشم از ارگان هاي پيچيده آسانتر است. به اين خاطر كه اين تغييرات را مي تواند به عنوان تعديلات منطقه اي كمي نسبت به يك بافت و پيش ساخته ديد.
به منظور تعيين تعداد توليدات لازم براي تكامل يك چشم، نيلسون به سادگي طرح محاسباتي ترتيب منطقي تغييراتي كه از يك موضع حساس به نور منجر به عدسي فوق پيشرفتة چشمي مي شود را ساخت.
نيلسون يك ارگانيسم با سلول هاي حساس به روشنايي كه روي يك پيش زمينة تاريك ساكن اند و براي تشخيص فضايي در يك گزينش قرار گرفته اند، فرض كرد. اولين روش براي توليد تشخيص فضايي فشار به وسط تكه و يا فشار روي لبه هاي تكه براي انقباض و بلند شدن است. اين تقعر اجازه مي دهد كه همبستگي مبهم نور در جايي كه نمايش يك منطقه روي تكه به زاوية منبع روشنايي وابسته است، جا بگيرند.
اين تكامل تقعر بايد براي شكل گيري يك فشار در تكه، نسبت به انقباض يك روزنه از طريق بلند شدن و انقباض غشاي احاطه كنندة رنگدانه، مطلوب باشد. اين منجر به فرورفتگي در چشم مي شود كه مشابه چشم بعضي از نرم تنان است.
اين pinhole – like براي وضوح جزئيات خوب نيستند و تصاوير بسيار تاريكي خلق مي كنند. به خاطر همين، هر گونه تغييرات كه شفافيت و آشكار سازي را بهبود بخشد، مطلوب خواهد بود.
دو راه براي اين تغييرات، توسعة يك عدسي و يا افزايش اندازة چشم است. اگر چه افزایش اندازه چشم مشکلات جسمانی را پیش رو دارد و تیزبینی کمتری نسبت به عدسی خواهد داشت.
انواع چشم:
چشم معمولي سه نوع است:
1- لنز ساده يا چشم دوربين: در مهره داران و برخي از بي مهرگان يافت مي شود. اين نوع از چشم را انسان داراست. حتي شما مي توانيد همين الان با انگشتتان به آن فشار وارد كنيد. ادامه بدهيد، آرام . همينه.
2- چشم آينه اي مقعر: در حلزون دو كپه اي يا صدف خوراكي از جنس pecten و تعداد كمي سخت پوستان دو كپه اي و خرچمگ ها يافت مي شود. اين نوع چشم براي جانور توليد روشنايي و تصاوير مبهم مي كند.
3- The pinhale Eye: چشم هاي حفره دار يا سوراخ عمدتاً در نرم تنان يافت مي شود و فقطمي توانند مكان اشياء را تشخيص دهند.
چشم مركب:
كه از دريافت كننده هاي چشم هاي سادة حشرات تشكيل شده و هر كداك به عنوان يك دريافت كننده (رسپنتور) بينايي جداگانه عمل مي كنند.
چشم متضاد: چشم هاي سادة حشرات به طور مستقل عمل مي كنند. (غيروابسته)
The superposition Eye: چشم هاي سادة حشرات براي توليد تصوير بزرگتر و با جزئيات بيشتر بر روي شبكيه با هم همكاري مي كنند.
دو نوع رسپنتورهاي حساس به نور مرئي:
همة رسپنتورهاي حساس به نور از يك رنگدانة حساس به نور كه از ويتاكين A تركيب شده با opsin مشتق شده است، استفاده مي كنند. بعد از اين كه رنگدانة نوري در معرض روشنايي قرار گرفت، opsin به يك G – پروتئين (نروترانسميتر متداول) پيوند مي خورد. اين مشابهت ها يك ريشه مشترك را پيشنهاد مي كند.
گيرنده هاي نوري: Rhabdomeric (در protosomes) كه عمدتاً در چشمان بندپايان يافت مي شود.
گيرنده هاي نوري مژك دار (در deuterosomes) در مهره داران مشترك است.
Rhabdomeric photoreceptors :
مناطق سطحي خود را به وسيلة بلند كردن سطوح قله اي در چين هاي بسيار زياد، افزايش مي دهد. (نظرية موي بارت سيمپسون). ريخت شناسي هاي بسياري براي اين گيرنده موجود است.
گيرنده هاي نوري مژك دار:
مناطق سطحي غشاء را به وسيلة تعديل مژك ها افزايش مي دهد. غشاي مژك دار متسع شده و وارد چين هاي عميق مي شود. در نتيجه ناحية واقعي گيرندة سلول مانند بسته هاي ديسك مي شود.
ميله ها و مخروط ها به نظر مي رسد كه از مواد متشكله متداول گيرندة نوري مژك دار منشأ گرفته و استنتاج شده اند، در حالي كه عقدة شبكيه اي، amacrine و سلول هاي افقي به نظر مي رسد كه از يك ماده متشكله Rhobdonieric استنتاج شده است.
چشم ها چگونه تكامل يافته اند؟
متخصصان افرينش اغلب از چشم به عنوان يك نقطه بحث و مناظره استفاده مي كنند تا نشان دهند كه تكامل نقض شده است و ذكر كرده اند كه چشم خيلي پيچيده است و از نظر تكاملي عاليتر از تكامل يافته است. خود «داروين» نوشته است كه «براي فرض كردن اين كه چشم مي تواند به وسيلة گزينش هاي طبيعي تشكيل شده باشد، به نظر مي رسد، من آزادانه اعتراف مي كنم كه اين بحث نامعقولي در بالاترين حد ممكن است. او در ادامه به هر حال، با تأكيد گفت: چشم ها احتمالاً از نرون هاي حساس به روشنايي تكامل يافته اند، به نظر مي رسد كه واقعيت باشد.
تكامل چشم:
تحقيقات دَن اِ نيلسون و سوزان بِلگِر نشان مي دهد كه در حقيقت تخمين تعداد توليدات ضروري براي تكامل يك چشم از ارگان هاي پيچيده آسانتر است. به اين خاطر كه اين تغييرات را مي تواند به عنوان تعديلات منطقه اي كمي نسبت به يك بافت و پيش ساخته ديد.
به منظور تعيين تعداد توليدات لازم براي تكامل يك چشم، نيلسون به سادگي طرح محاسباتي ترتيب منطقي تغييراتي كه از يك موضع حساس به نور منجر به عدسي فوق پيشرفتة چشمي مي شود را ساخت.
نيلسون يك ارگانيسم با سلول هاي حساس به روشنايي كه روي يك پيش زمينة تاريك ساكن اند و براي تشخيص فضايي در يك گزينش قرار گرفته اند، فرض كرد. اولين روش براي توليد تشخيص فضايي فشار به وسط تكه و يا فشار روي لبه هاي تكه براي انقباض و بلند شدن است. اين تقعر اجازه مي دهد كه همبستگي مبهم نور در جايي كه نمايش يك منطقه روي تكه به زاوية منبع روشنايي وابسته است، جا بگيرند.
اين تكامل تقعر بايد براي شكل گيري يك فشار در تكه، نسبت به انقباض يك روزنه از طريق بلند شدن و انقباض غشاي احاطه كنندة رنگدانه، مطلوب باشد. اين منجر به فرورفتگي در چشم مي شود كه مشابه چشم بعضي از نرم تنان است.
اين pinhole – like براي وضوح جزئيات خوب نيستند و تصاوير بسيار تاريكي خلق مي كنند. به خاطر همين، هر گونه تغييرات كه شفافيت و آشكار سازي را بهبود بخشد، مطلوب خواهد بود.
دو راه براي اين تغييرات، توسعة يك عدسي و يا افزايش اندازة چشم است. اگر چه افزایش اندازه چشم مشکلات جسمانی را پیش رو دارد و تیزبینی کمتری نسبت به عدسی خواهد داشت.
اشتراک در:
پستها (Atom)