





An extracellular matrix molecule that is selectively expressed during development is important for gastrulation in the sea urchin embryo.
Berg LK, Chen SW, Wessel GM.
Department of Molecular and Cell Biology & Biochemistry, Brown University, Providence, RI 02912, USA.
The extracellular matrix is important in the regulation of many cellular events of early development including migration, shape change, proliferation and gene expression. In the sea urchin embryo, disruption of the extracellular matrix results in selective defects in each of these events during gastrulation. Here we describe a new molecule of the extracellular matrix in Lytechinus variegatus, referred to as ECM 18, that has several important features. First, antibody interference of ECM 18 results in a profound but reversible inhibition of primary mesenchyme cell organization and endoderm morphogenesis during gastrulation. Second, during gastrulation, ECM 18 mRNA accumulates to highest levels in the invaginating endoderm and the ECM 18 protein deposited in the basal lamina surrounding the archenteron as well as in other areas of the blastocoel wall. Immunolocalization by fluorescence and electron microscopy demonstrates the selective accumulation of ECM 18 in the extracellular matrix. Third, although the mRNA encoding ECM 18 is present throughout development, the protein accumulates only during gastrulation. ECM 18 protein is not detected in eggs or early embryos and analysis of polysome-associated mRNA suggests that at least part of the translational regulation of ECM 18 is at the level of ECM 18 mRNA-polysome formation. Finally, sequence analysis of ECM 18 shows that the protein contains a repeat sequence with a conserved cysteine motif, suggestive of involvement in protein-protein interactions. Thus, ECM 18 appears to be important in mediating select morphogenetic changes during gastrulation and the pattern of its expression in the embryo is unique among the extracellular matrix molecules known in this embryo.
PMID: 8625821 [PubMed - indexed for MEDLINE]
transcription Factor Activation in the Sea Urchin Embryo
It should be noted that the biochemical nature of the vegetalizing signal(s) has not yet been elucidated. Hörstadius interpreted these experiments in terms of gradients of animalizing and vegetalizing substances. More recently, Davidson (1989) sought to explain these phenomena in terms of transcriptional regulatory proteins (such as those that bind to promoters or enhancers) that are localized throughout the egg and that become activated by inductive interactions between adjacent cells. One of these factors, the one in the micromere precursors of the primary mesenchyme cells, is thought to become active autonomously, early in cleavage. This factor would cause the determination of the micromeres, alter their cell membranes so they could react with the cells above them, and initiate the cascade of determination. By the end of blastulation, there would be five territories that would constitute both the precursor cells for certain regions of the pluteus larva and the regions of specific gene activation. For example, the cytoskeletal actin gene, which is expressed only in the aboral ectoderm cells, cannot be activated anywhere else in the embryo. Similarly, if one injects the gene for the skeletal matrix protein anywhere else in the embryo except the region of skeletogenic mesenchyme, it will not be activated (Hough-Evans et al., 1987; Sucov et al., 1988). Therefore, each territory would constitute a region of differential gene expression that would be reflected in different parts of the organism being formed.
However, if the embryo is perturbed, the neighbors would differ. New fates would ensue as the new vegetal cells activated their skeletogenic mesenchyme determinants and started activation of their neighbors. To see if this was indeed the case, Ransick and Davidson (1993) transplanted micromeres into the animal cap of an 8-cell sea urchin embryo. These skeletogenic mesenchyme precursor cells induced the formation of the archenteron and the expression of the archenteron-specific genes. The fluorescently labeled micromeres did not contribute structurally to the secondary archenteron, but provided a short-range inductive signal that respecified the animal cap cells.
Literature Cited
Davidson, E. H. 1989. Lineage-specific gene expression and the regulative capacities of the sea urchin embryo: a proposed mechanism. Development 105: 421-446.
Hough-Evans, B. R., Franks, R. R., Cameron, R. A., Britten, R. J. and Davidson, E. H. 1987. Correct cell type-specific expression of a fusion gene injected into sea urchin eggs. Dev. Biol. 121: 576-579.
Ransick, A. and Davidson, E. H. 1993. A complete second gut induced by transplanted micromeres in the sea urchin embryo. Science 259: 1134-1138.
Sucov, H. M., Hough-Evans, B. R., Franks, R. R., Britten, R. J. and Davidson, E. H. 1988. A regulatory domain that directs lineage-specific expression of a skeletal matrix protein in the sea urchin embryo. Genes Dev. 2: 1238-1250.
© All the material on this website is protected by copyright. It may not be reproduced in any form without permission from the copyright holder.
HOME :: CHAPTER 8 :: SEA URCHIN CELL SPECIFICATION :: TRANSCRIPTION FACTOR ACTIVATION IN THE SEA URCHIN EMBRYO PREVIOUS :: NEXT
Home Link
گاسترو لاسون درتوتیای دریایی
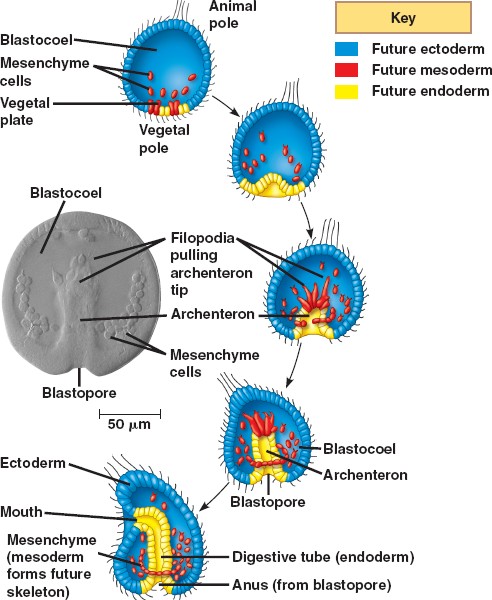
توضیحات زیر مربوط به گاسترولاسیون در خارپوستان است .(جاندارانی با 3 لایه ی جنینی) هرچند توضیحات مربوط به تصویر گاسترلاسیون جانورانی است با دو لایه ی جنینی.
توتياي دریایی تسهیم کامل داردتسهیم اول ودوم هر دو نصف النهاری و عمود بر هم می باشد. تسهیم سوم مداری وعمود بردو صفحه ی تقسیم اول ودوم انجام شده قطب جانوری وگیاهی را از هم جدا می کند تسهیم چهارم کاملا متفاوت از سه تسهیم اول می باشد درتوتیای دریایی از کلیواژ ساده به طرف شکافتگی چهارم (پیشرفته تر ) هدایت شده ، چهار بلاستومر قطب نباتی به طور نامساوی تقسیم شده ، 4 میکرومر را در نزدیکی قطب نباتی و 4 ماکرومر را در بخش میانی ایجاد کرده سلول های قطب حیوانی نیز تقسیم نصف النهاری انجام داده و مزومرها را تولید می کند .
مرحله ی بلاستولای تکوین در توتیای در یایی از مرحله 128 سلولی شروع می شود.در این زمان سلول ها کرهی تو خالی را بوجود می آورند که حفره مرکزی بلاستوسل را در برمی گیرد.که حاوی مایع پروتیینی می باشد.بلاستومرها اطراف این حفره با اتصالاتی محکم صفحه اپی تلیالی پیوسته ای را ایجاد می کند.همچنین هر یک از سلولها (بلاستومرها)دربخشی از غشای سلولها که دور از حفره مرکزی بلاستوسل قرار دارد مژه دار می شود و بلاستولای مژه دار در پوشش لقاحی خود شروع به چرخیدن کرده ومتعاقب آن تمایز می یابند.
جنین در قطب نباتی شروع به پهن شدن کرده و به شکل پلاک نباتی در می آید ،برخی از سلول ها از قطب نباتی مهاجرت کرده وپس ازعبورتبدیل به سلولهای مزانشیمی اولیه می شوند ،سلول های مزانشیمی اولیه تقسیم شده و در امتداد ماتریکس خارج سلولی به طرف بخش های مختلف بلاستوسل می روند .
این مهاجرت بنابر اعتقادی بستگی دارد به پروتئوگلیکان سطح سلول ها و ملکول هایی بر روی غشای پایه که فیبرونکتین نامیده می شوند . حرکت سلول ها توسط فیلوپودیاها هدایت می شود به عبارتی این فیلوپودیاها محل سلول هدف را می شناسند.
ساختار (گودالی)به نام آرکنترون را ایجاد می کند که در دهانه ی آن بلاستوپور نامیده می شود .
توسعه آرکنترون توسط 3 مکانیسم ادامه می یابد:
1)درون خزیدگی اولیه ایجاد می شود . با انبساط پی در پی لایه ی درونی ساخته شده از فیبروپیلن و لایه ی خارجی ساخته شده از هیالین.
2)آرکنترون شکل می گیرد از توسعه ی همگرا (توسعه ای که به همه ی قسمت ها برسد ) . توسعه ی همگرا زمانی اتفاق می افتد که سلول ها در میان بافت باریکی جای گرفته و این ساختار به جلو حرکت می کند .
3)مزانشیم ثانویه کشیده می شود به نوک آرکنترون . مزانشیم ثانویه شکل می گیرد از سلول هایی که وارد می شوند و متصل به سقف آرکنترون باقی می مانند .
فیلوپودیا راهنمای هدایت است . که بعداً این ساختار در ناحیه دهان یافت می شود به محض رسیدن به هدف فیلوپودیا با اکتودرم تماس می یابد و سلول مزانشیم به قطب حیوانی می رسد و در نهایت یک حفره ی گوارشی ایجاد می شود که از همه ی راههای میان
جنین می گذرد .
4)لایه های 3 گانه ی جنینی شکل می گیرد . اندودرم به قطعه ی گوارشی ، اکتودرم شامل سلول های خارج گاسترولاست که در گاسترولاسیون نقش کمی ایفا میکند واین اکتودرم به پوست و سیستم عصبی مرکزی توسعه می یابد، مزودرم شامل سلول های مزانشیمی است که در بلاستوسل توسعه می یابد و تبدیل به اندام داخلی جنین می شود
مهاجرت انفرادی مزانشیم اولیه
عملکرد سلولهای مزانشیمی اولیه .مدت زمان کوتاهی پس از خروج بلاستومرها از پوشش لقاحی گروهی از سلولهای مشتق شده از میکرومرها نوعی تغیر شکل از حالت اپی تلیا ل به مزانشیمی پیدا می کنند.این سلولها اسکلت سلولی خود را تعغیر داده و بطری شکل می شوند همچنین اتصال خودرابا سلولهای جانبی از دست داده و با جدا شدن از لایه راسی وارد بلاستوسل می شوند.(شکل15-8ساعت 10-9)
این سلولهای مشتق شده از میکرومرها مزانشیم اولیه نامیده می شوند.از انجایی که این سلولها اسکلت لا روی راتشکیل میدهند و مزانشیم اسکلت زا نیز گفته می شود این سلولهای مزانشیمی قادرند زائدهایی بلند و نازک و منقبض شونده (به نام فیلو پودیا را به وجود اورند.
.در ابتدا به نظر می رسد که این سلولها به صورت تصادفی در امتداد سطح درونی بلا ستوسل حرکت می کنند.برای انجام حرکت زوائد فیلو پود یایی این سلو لها به طور متناوب اتصالاتی را با دیواره ی بلاستوسل برقرار کرده وسپس جدا می شوند نهایتا این سلولها در ناحیه شکمی – جانبی اینده ی بلاستوسل مستقر میگردند.در این مکان به طناب سین سیشیومی که در تشکیل محورکربنات کلسیمی اسپیکول های اسکلتی لارو نقش دارد ملحق می شوند (شکل d-e14-8)
شكل 8-15
اهمیت لایه ی ماده ی زمینه ای برون سلولی درون بلا ستوسل.
مهاجرت نسل میکرمری به درون بلا ستوسل در نتیجه ی از دست رفتن تمایل این سلولهای مزانشیمی اولیه برای اتصال به سلولهای مجاور و غشای هیالینی ودر عوض تمایل بالای انها برای اتصال یافتن به گروهی از پروتئینها که دربلاستوسل قرار گرفته اند رخ می دهد این نوع مدل مهاجرت مزانشیمی برای اولین بارتوسطgustafsonوwolpertدر 1967 ارائه شد ودر سال 1985زمانی که rachelfinkوdavidmcclay میزان قدرت اتصال بلا ستومرهای توتیایی دریایی به لایه ی هیا لینی غشای پایه ی بلاستوسل وسایر بلاستومرها رااندازه گیری کردند تایید گردید.
تمامی سلولهای بلاستولا در ابتدا از سطح بیرونی خود به لایه ی هیالینی و از سطح درونی خود به غشای پایه ی ترشح شده توسط خودشان متصل هستند .هر سلول از سطوح جانبی نیز به سلولهای همسایه اتصال دارد.finkوmcclayدریافتند که سلولهای اکتودرمی و اندودرمی اینده ی جنین (به ترتیب اعقاب مزومرها وماکرومرها)محکم به یکدیگر و به لایه ی هیالینی متصل می شوند اما اتصال این سلولها به غشای پایه سست است میکرومها نیز در ابتدا الگوی اتصالی مشابهی را دارند اما الگوی انها حین گاسترو لاسیون تغییر می کند در حالی که سلولهای دیگر اتصال محکم خود را با لایه ی هیالینی و سلولهای مجاور حفظ می کنند پیش سازهای مزانشیمی اولیه تمایل خود را نسبت به این ساختارها از دست می دهند (این تمایل به حدود 2درصد تمایل اولیه می رسد)اما تمایل انها نسبت به ترکیبات موجود درغشا ی پایه و ماده ی زمینه ای برون سلولی (نظیر فیبرونکتین)صد برابر بیشتر می شود
تمامی سلولهای بلاستولا در ابتدا از سطح بیرونی خود به لایه ی هیالینی و از سطح درونی خود به غشای پایه ی ترشح شده توسط خودشان متصل هستند .هر سلول از سطوح جانبی نیز به سلولهای همسایه اتصال دارد.finkوmcclayدریافتند که سلولهای اکتودرمی و اندودرمی اینده ی جنین (به ترتیب اعقاب مزومرها وماکرومرها)محکم به یکدیگر و به لایه ی هیالینی متصل می شوند اما اتصال این سلولها به غشای پایه سست است میکرومها نیز در ابتدا الگوی اتصالی مشابهی را دارند اما الگوی انها حین گاسترو لاسیون تغییر می کند در حالی که سلولهای دیگر اتصال محکم خود را با لایه ی هیالینی و سلولهای مجاور حفظ می کنند پیش سازهای مزانشیمی اولیه تمایل خود را نسبت به این ساختارها از دست می دهند (این تمایل به حدود 2درصد تمایل اولیه می رسد)اما تمایل انها نسبت به ترکیبات موجود درغشا ی پایه و ماده ی زمینه ای برون سلولی (نظیر فیبرونکتین)صد برابر بیشتر می شود
این تغییر در تمایلات سلولی موجب ازادسازی اتصالات سلولی میکرومرها از لایه ی هیالینی خارجی و سلولهای مجاور شده و باعث می شود که از طریق غشای پایه به داخل کشیده شده وبه درون بلاستوسل مهاجرت کنند (شکل a 16-8) این تغییر در تمایلات به دلیل تغییر در مولکولهای سطح سلولی است که در این زمان ایجاد می شودهمچنین پرو تئین هایی نظیر فیبرونکتین اینتگرین لامینین1وکادهرین ها نیز در این مهاجرت سلولی شرکت دارند .
همان گونه که درشکل cوb16-8نشان داده شده غلظت بالایی از مواد برون سلولی در اطراف سلولهای مزانشیمی اولیه ی در حال مهاجرت دیده می شود.در داخل بلاستوسل به نظر می رسید که سلولهای مزانشیمی اولیه در امتداد ماده زمینه ی برون سلولی دیواره بلاستوسل مهاجرت کرده و فیلوپودیای خود رابه سمت جلو گسترده می کنندچندین نوع پروتئین(نظیر فیبرونکتین و نوع خاصی گلیکوپروتئین سولفاته)
در شروع و حفظ این مهاجرت لازم و ضروری هستنداما این علایم جهت یابی کافی نیستند چرا که سلولهای در مهاجرت((میدانند))که چه موقع حرکت خود را متوقف کنند و اسپیکولها را نزدیک استوای بلاستوسل تشكيل دهند.
سلولهای مزانشیم اولیه به صورت یک حلقه در موقعیتی خاص و در امتداد محور جانوری-نباتی سازمان دهی میشوند-بسیاری از سلولهای مزانشیم اولیه در دو ناحیه نزدیک بخش شکمی اینده ی لارو تجمع
یافته با یکدیگر ترکیب شده و شروع به تشکیل اسپیکول می کنند(شکل17-8)
شكل 8-17
اگر میکرومر نشان دار شده ای از یک جنین به بلاستوسل جنین دیگری در مرحله ی گاسترولاسیون تزریق شود به محل صحیح مهاجرت کرده ودر تشکیل اسپیکول های جنینی شرکت می کندتصور می شود که اطلاعات مکانی مورد نیاز توسط سلولهای اکتودرمی اینده وغشا ی پایه ی انه فراهم می شود (شکلa18-8}
تنها سلولهای مزانشیمی اولیه (ونه انواع دیگر سلولها)قادرند به این علایم الگوبندی پاسخ دهند
شكل 8-18
.
millerوهمکارانش در 1995از وجود فیلوپودیا کاملا واضح (با قطر3/0ميكرو متر)در سلولهای مزانشیمی تشکیل دهنده ی اسکلت سلولی خبر دادند
به نظر می رسد که این فیلو پودیا در حرکت نقش فعالی نداشته بلکه به جستجوی دیواره ی بلاستوسل می پردازد وشاید در پاسخگویی به علایم الگو بندی پشتی – شکمی و جانوری نباتی صادر شده از اکتو درم نقش داشته باشد.
مرحله ی اول درون روی آرکنترون
هم زمان که سلول های مزانشیمی اولیه ی قطب نباتی جنین کروی را ترک می کنند ، تغییرات مهمی در سلول های باقی مانده ایجاد می شود.سلول های باقی مانده ضخیم و پهن شده و صفحه ی نباتی را تشکيل دهند.در نتیجه شکل جنین تغییر می کند شكل8 -51)ساعت 9سلول های صفحه ی نباتی به یکدیگر و به لایه ی هیالینی سلول تخم متصل باقی مانده و به گونه ایی حرکت میکنند تا فضاهای خالی به وجود آمده ی حاصل از مهاجرت مزانشیم اولیه را پر کنند. به علاوه صفحه نباتی به درون خم شده وپس از درون روی حدود 4/1الی 2/1به طور ناگهانی متوقف می شوند دهانه این ناحیه درون رفته (آرکنترون ) بلاستو پور نامیده میشود.
بررسیها نشان داده که لایه هیالینی موجوددر صفحه نباتی به دلیل تغییر در ترکیباتش به سمت درون خمیده میشود.لایه هیالینی درواقع از دو تیغه تشکیل شده است یک تیغه بیرونی متشکل از پروتیین هیالین و یک پروتیین درونی متشکل از پروتیین های فیبروپیلین .فیبروپیلین ها در گرانول های ترشحی تخمک ذخیره شده وپس از اگزوسیتوز گرانول های قشری ترشح می شوند و منجر به آزاد سازی پروتیین هیالین می گردند.درمرحله بلاستولا فیبروپلین ها شبکه ای تور ماننددر اطراف جنین ایجاد میکند .در حین درون روی سلول های صفحه نباتی شرو به ترشح گلیکو پروتیین کندرویتین سولفات به درون تیغه درونی لایه هیالینی میکند در نتیجهناحیه نباتی لایه هیالینی خمیده میشود اندکی بعد یک نیروی ثانویه که از حرکت سلولهای اپی تلیالی مجاور صفحه نباتی حاصل میشود با کشیدن لایه خمیده به درون موجب تسهیل در فرآیند درون روی می گردد.
سلول های مزانشیم ثانویه اولین گروه از سلول هایی هستند که درون روی نموده ، به درون بلاستوسل پیش روی می کنند و راس آرکنترون را تشکیل می دهند . آنها نهایتاً سلول های رنگدانه دار ، عضلات اطراف لوله ی گوارشی ، و کیسه های سلومی را تشکیل
می دهند سلول های اندودرمی مجاور مزانشیم مشتق شده از میکرومر ،لوله ی گوارشی قدامی را به وجود می آورند که فاصله ی بیشتری را در بلاستوسل مهاجرت می کند . لایه ی بعد سلول های اندودرمی لوله ی گوارشی میانی را به وجود می آورد . ردیف محیطی آخر نیز درون روی نموده ، لوله ی گوارش خلفی و مخرج را تشکیل می دهد
مراحل دوم و سوم درون روی آرکنترون
دنبال درون روی اول ، فاز دوم شکل گیری آرکنترون شروع می شود . طی این مرحله آرکنترون گسترده شده به گونه ایی که حتی ممکن است طول آن سه برابر شود . در طی این گسترده
شدن لوله ی گوارش کوتاه و عریض به لوله ای باریک و طویل تغییر شکل می دهد . برای این طریق مهاجرت بر روی یکدیگر و پهن شدن باز آرایی می شوند .
پدیده ایی که طی آن سلول ها در هم فرو می روند و موجب نازک شدن بافت شده و هم زمان یه سمت جلو حرکت می کنند بسط همگرا نامیده میشود .
در تمام گو نه های توتیای در یایی مرحله سوم طویل شدن آرکنترون نیز دیده می شود این مرحله انتهایی در اثر نیروی کششی که توسط سلولهای مزانشیم ثانویه ایجاد میشود آغاز میگردد .این سلولها راس آرکنترون را ساخته ودر همان جا مستقر میشوند سلولهای مذکوردر مایع بلاستوسل زواید فیلوپودیایی ایجاد می کنند که به طرف سطح درونی دیواره بلاستوسل کشیده میشود.فیلوپودیاهادرناحیه اتصالی بین بلاستومرهابه دیواره متصل شده سپس با کوتاه شدن خود آرکنترون را به بالا میکشند(شکل های 8-15و 8-21 ساعت 12و13)
. اگرتعدادی از سلول های مزانشیم ثانویه حذف شوند طویل شدن ادمه می یابد، اگرچه سرعت آن کند می شود . در این گونه سلول های مزانشیم ثانویه نقشی اساسی در کشیده شدن آرکنترون به سمت دیوار بلاستوسل حین مرحله ی انتهایی درون روی بازی می کنند. اما آیا فیلوپودیای مزانشیم ثانویه می تواند به هر بخشی از دیواره ی بلاستوسل متصل شود، یا این که هدف خاصی در نیم کره جانوری باید وجود داشته باشد تا اتصال صورت گیرد؟ آیا بخشی از دیواره ی بلاستوسل وجود دارد که متعهد شده تا بخش شکمی لارورابه وجود آورد؟ مطالعات صورت گرفته توسط Hardin وMcCIay در1990 نشان می دهد که ناحیه ی هدف ویژه ای برای فیلوپود یا وجود دارد که از سایر نواحی قطب جانوری متفاوت است فیلوپود یاگسترش می یابد و به صورت تصادفی به دیواره ی بلاستوسل متصل شده وسپس آزاد می گردد. با این وجودزمانی که فیلوپود یا با ناحیه ی خاصی از دیواره برخورد می کند اتصال خود را حفظ کرده ، منقبض شده وآرکنترون را به سمت خود می کشد. زمانی که Hardin وMcCIay سمت دیگر دیواره ی بلاستوسل رادرمعرض فیلوپود یاها قراردادند، فیلوپود یا به گسترش خود ادامه داده وبعد از برخورد با این دیواره ی دست کاری شده آن را رها نمودند. تنها زمانی که فیلوپود یا بافت هدف مورد نظر را پیدا کرد، حرکات خود را متوقف نمود
. اگرگاسترولا به اندازه ای طویل می شد که فیلوپود یا هرگزنمی توانست به ناحیه ی هدف برسد، سلول های مزانشیم ثانویه به جستجوی خود ادمه می دادند وسرانجام آرکنترون را رها کرده وبه صورت آزاد، هدف خود را جستجو می کردند. بنابراین،آنچه واضح است این است که در ناحیه ای که قرار است بخش شکمی لارورا به وجود آورد وهمچنین ناحیه ای از آکنترون که دهان آینده ی جنین را به وجودخواهد آورد،
بخش هدفی وجود دارد که توسط سلول های مزانشیم ثانویه شناسایی می شود . هم زمان با برخورد راُس آرکنترون با ناحیه ی هدف دیواره ی بلا ستوسل سلول های مزانشیم ثانویه روی بلاستوسل پخش می شوند، سپس تکثیر شده واندام های مزودرمی را به وجود می آورند (شکل 15-8 ساعت 5/13). جایی که آرکنترون با دیواربرخورد می کند، نهایتاً به دهان موجود تبدیل خواهد شد
سلول ها مزانشیمی اسکلت زاوارد rudiment شده تا صفحات اسکلتی اولیه سند شوند سمت چپ پلوتئوس سطح دهانی آینده ی جنین توتیای دریایی بالا را به وجود می آورد، حین دگردیسی، imaginal rudime لاروجدا شده سپس لاروتخریب می شود،درحالی که naginal rudiment ( که حالا جوانه نامیده می شود) مجد داً لوله ی گوارش خود ایجاد کرده ودر کف اقیانوس قرار می گیرد. همچنان به مواد غذایی که لاروها شده دریافت کرده وابسته است. الگوی گاسترولاسیون خاربوستان نمونه ی تکاملی تکوین پس دهانیازبه
حساب می آید.درپس دهانیان (خارپوستان ، تونیکات ها سفالوکوردات ها و مهره داران) منفذ اولیه تبدیل به مخرج می شود، درحالیکه منفذ دوم دهان را تشکیل می دهد (دوتروستوم به معنی دهان ثانویه). به علاوه درتوتیای
دریایی پدپده ی گسترش همگرا واستفاده ازبیان ژن Nodal درتثبیت محورها دیده می شود
Berg LK, Chen SW, Wessel GM.
Department of Molecular and Cell Biology & Biochemistry, Brown University, Providence, RI 02912, USA.
The extracellular matrix is important in the regulation of many cellular events of early development including migration, shape change, proliferation and gene expression. In the sea urchin embryo, disruption of the extracellular matrix results in selective defects in each of these events during gastrulation. Here we describe a new molecule of the extracellular matrix in Lytechinus variegatus, referred to as ECM 18, that has several important features. First, antibody interference of ECM 18 results in a profound but reversible inhibition of primary mesenchyme cell organization and endoderm morphogenesis during gastrulation. Second, during gastrulation, ECM 18 mRNA accumulates to highest levels in the invaginating endoderm and the ECM 18 protein deposited in the basal lamina surrounding the archenteron as well as in other areas of the blastocoel wall. Immunolocalization by fluorescence and electron microscopy demonstrates the selective accumulation of ECM 18 in the extracellular matrix. Third, although the mRNA encoding ECM 18 is present throughout development, the protein accumulates only during gastrulation. ECM 18 protein is not detected in eggs or early embryos and analysis of polysome-associated mRNA suggests that at least part of the translational regulation of ECM 18 is at the level of ECM 18 mRNA-polysome formation. Finally, sequence analysis of ECM 18 shows that the protein contains a repeat sequence with a conserved cysteine motif, suggestive of involvement in protein-protein interactions. Thus, ECM 18 appears to be important in mediating select morphogenetic changes during gastrulation and the pattern of its expression in the embryo is unique among the extracellular matrix molecules known in this embryo.
PMID: 8625821 [PubMed - indexed for MEDLINE]
transcription Factor Activation in the Sea Urchin Embryo
It should be noted that the biochemical nature of the vegetalizing signal(s) has not yet been elucidated. Hörstadius interpreted these experiments in terms of gradients of animalizing and vegetalizing substances. More recently, Davidson (1989) sought to explain these phenomena in terms of transcriptional regulatory proteins (such as those that bind to promoters or enhancers) that are localized throughout the egg and that become activated by inductive interactions between adjacent cells. One of these factors, the one in the micromere precursors of the primary mesenchyme cells, is thought to become active autonomously, early in cleavage. This factor would cause the determination of the micromeres, alter their cell membranes so they could react with the cells above them, and initiate the cascade of determination. By the end of blastulation, there would be five territories that would constitute both the precursor cells for certain regions of the pluteus larva and the regions of specific gene activation. For example, the cytoskeletal actin gene, which is expressed only in the aboral ectoderm cells, cannot be activated anywhere else in the embryo. Similarly, if one injects the gene for the skeletal matrix protein anywhere else in the embryo except the region of skeletogenic mesenchyme, it will not be activated (Hough-Evans et al., 1987; Sucov et al., 1988). Therefore, each territory would constitute a region of differential gene expression that would be reflected in different parts of the organism being formed.
However, if the embryo is perturbed, the neighbors would differ. New fates would ensue as the new vegetal cells activated their skeletogenic mesenchyme determinants and started activation of their neighbors. To see if this was indeed the case, Ransick and Davidson (1993) transplanted micromeres into the animal cap of an 8-cell sea urchin embryo. These skeletogenic mesenchyme precursor cells induced the formation of the archenteron and the expression of the archenteron-specific genes. The fluorescently labeled micromeres did not contribute structurally to the secondary archenteron, but provided a short-range inductive signal that respecified the animal cap cells.
Literature Cited
Davidson, E. H. 1989. Lineage-specific gene expression and the regulative capacities of the sea urchin embryo: a proposed mechanism. Development 105: 421-446.
Hough-Evans, B. R., Franks, R. R., Cameron, R. A., Britten, R. J. and Davidson, E. H. 1987. Correct cell type-specific expression of a fusion gene injected into sea urchin eggs. Dev. Biol. 121: 576-579.
Ransick, A. and Davidson, E. H. 1993. A complete second gut induced by transplanted micromeres in the sea urchin embryo. Science 259: 1134-1138.
Sucov, H. M., Hough-Evans, B. R., Franks, R. R., Britten, R. J. and Davidson, E. H. 1988. A regulatory domain that directs lineage-specific expression of a skeletal matrix protein in the sea urchin embryo. Genes Dev. 2: 1238-1250.
© All the material on this website is protected by copyright. It may not be reproduced in any form without permission from the copyright holder.
HOME :: CHAPTER 8 :: SEA URCHIN CELL SPECIFICATION :: TRANSCRIPTION FACTOR ACTIVATION IN THE SEA URCHIN EMBRYO PREVIOUS :: NEXT
Home Link
گاسترو لاسون درتوتیای دریایی
توضیحات زیر مربوط به گاسترولاسیون در خارپوستان است .(جاندارانی با 3 لایه ی جنینی) هرچند توضیحات مربوط به تصویر گاسترلاسیون جانورانی است با دو لایه ی جنینی.
توتياي دریایی تسهیم کامل داردتسهیم اول ودوم هر دو نصف النهاری و عمود بر هم می باشد. تسهیم سوم مداری وعمود بردو صفحه ی تقسیم اول ودوم انجام شده قطب جانوری وگیاهی را از هم جدا می کند تسهیم چهارم کاملا متفاوت از سه تسهیم اول می باشد درتوتیای دریایی از کلیواژ ساده به طرف شکافتگی چهارم (پیشرفته تر ) هدایت شده ، چهار بلاستومر قطب نباتی به طور نامساوی تقسیم شده ، 4 میکرومر را در نزدیکی قطب نباتی و 4 ماکرومر را در بخش میانی ایجاد کرده سلول های قطب حیوانی نیز تقسیم نصف النهاری انجام داده و مزومرها را تولید می کند .
مرحله ی بلاستولای تکوین در توتیای در یایی از مرحله 128 سلولی شروع می شود.در این زمان سلول ها کرهی تو خالی را بوجود می آورند که حفره مرکزی بلاستوسل را در برمی گیرد.که حاوی مایع پروتیینی می باشد.بلاستومرها اطراف این حفره با اتصالاتی محکم صفحه اپی تلیالی پیوسته ای را ایجاد می کند.همچنین هر یک از سلولها (بلاستومرها)دربخشی از غشای سلولها که دور از حفره مرکزی بلاستوسل قرار دارد مژه دار می شود و بلاستولای مژه دار در پوشش لقاحی خود شروع به چرخیدن کرده ومتعاقب آن تمایز می یابند.
جنین در قطب نباتی شروع به پهن شدن کرده و به شکل پلاک نباتی در می آید ،برخی از سلول ها از قطب نباتی مهاجرت کرده وپس ازعبورتبدیل به سلولهای مزانشیمی اولیه می شوند ،سلول های مزانشیمی اولیه تقسیم شده و در امتداد ماتریکس خارج سلولی به طرف بخش های مختلف بلاستوسل می روند .
این مهاجرت بنابر اعتقادی بستگی دارد به پروتئوگلیکان سطح سلول ها و ملکول هایی بر روی غشای پایه که فیبرونکتین نامیده می شوند . حرکت سلول ها توسط فیلوپودیاها هدایت می شود به عبارتی این فیلوپودیاها محل سلول هدف را می شناسند.
ساختار (گودالی)به نام آرکنترون را ایجاد می کند که در دهانه ی آن بلاستوپور نامیده می شود .
توسعه آرکنترون توسط 3 مکانیسم ادامه می یابد:
1)درون خزیدگی اولیه ایجاد می شود . با انبساط پی در پی لایه ی درونی ساخته شده از فیبروپیلن و لایه ی خارجی ساخته شده از هیالین.
2)آرکنترون شکل می گیرد از توسعه ی همگرا (توسعه ای که به همه ی قسمت ها برسد ) . توسعه ی همگرا زمانی اتفاق می افتد که سلول ها در میان بافت باریکی جای گرفته و این ساختار به جلو حرکت می کند .
3)مزانشیم ثانویه کشیده می شود به نوک آرکنترون . مزانشیم ثانویه شکل می گیرد از سلول هایی که وارد می شوند و متصل به سقف آرکنترون باقی می مانند .
فیلوپودیا راهنمای هدایت است . که بعداً این ساختار در ناحیه دهان یافت می شود به محض رسیدن به هدف فیلوپودیا با اکتودرم تماس می یابد و سلول مزانشیم به قطب حیوانی می رسد و در نهایت یک حفره ی گوارشی ایجاد می شود که از همه ی راههای میان
جنین می گذرد .
4)لایه های 3 گانه ی جنینی شکل می گیرد . اندودرم به قطعه ی گوارشی ، اکتودرم شامل سلول های خارج گاسترولاست که در گاسترولاسیون نقش کمی ایفا میکند واین اکتودرم به پوست و سیستم عصبی مرکزی توسعه می یابد، مزودرم شامل سلول های مزانشیمی است که در بلاستوسل توسعه می یابد و تبدیل به اندام داخلی جنین می شود
مهاجرت انفرادی مزانشیم اولیه
عملکرد سلولهای مزانشیمی اولیه .مدت زمان کوتاهی پس از خروج بلاستومرها از پوشش لقاحی گروهی از سلولهای مشتق شده از میکرومرها نوعی تغیر شکل از حالت اپی تلیا ل به مزانشیمی پیدا می کنند.این سلولها اسکلت سلولی خود را تعغیر داده و بطری شکل می شوند همچنین اتصال خودرابا سلولهای جانبی از دست داده و با جدا شدن از لایه راسی وارد بلاستوسل می شوند.(شکل15-8ساعت 10-9)
این سلولهای مشتق شده از میکرومرها مزانشیم اولیه نامیده می شوند.از انجایی که این سلولها اسکلت لا روی راتشکیل میدهند و مزانشیم اسکلت زا نیز گفته می شود این سلولهای مزانشیمی قادرند زائدهایی بلند و نازک و منقبض شونده (به نام فیلو پودیا را به وجود اورند.
.در ابتدا به نظر می رسد که این سلولها به صورت تصادفی در امتداد سطح درونی بلا ستوسل حرکت می کنند.برای انجام حرکت زوائد فیلو پود یایی این سلو لها به طور متناوب اتصالاتی را با دیواره ی بلاستوسل برقرار کرده وسپس جدا می شوند نهایتا این سلولها در ناحیه شکمی – جانبی اینده ی بلاستوسل مستقر میگردند.در این مکان به طناب سین سیشیومی که در تشکیل محورکربنات کلسیمی اسپیکول های اسکلتی لارو نقش دارد ملحق می شوند (شکل d-e14-8)
شكل 8-15
اهمیت لایه ی ماده ی زمینه ای برون سلولی درون بلا ستوسل.
مهاجرت نسل میکرمری به درون بلا ستوسل در نتیجه ی از دست رفتن تمایل این سلولهای مزانشیمی اولیه برای اتصال به سلولهای مجاور و غشای هیالینی ودر عوض تمایل بالای انها برای اتصال یافتن به گروهی از پروتئینها که دربلاستوسل قرار گرفته اند رخ می دهد این نوع مدل مهاجرت مزانشیمی برای اولین بارتوسطgustafsonوwolpertدر 1967 ارائه شد ودر سال 1985زمانی که rachelfinkوdavidmcclay میزان قدرت اتصال بلا ستومرهای توتیایی دریایی به لایه ی هیا لینی غشای پایه ی بلاستوسل وسایر بلاستومرها رااندازه گیری کردند تایید گردید.
تمامی سلولهای بلاستولا در ابتدا از سطح بیرونی خود به لایه ی هیالینی و از سطح درونی خود به غشای پایه ی ترشح شده توسط خودشان متصل هستند .هر سلول از سطوح جانبی نیز به سلولهای همسایه اتصال دارد.finkوmcclayدریافتند که سلولهای اکتودرمی و اندودرمی اینده ی جنین (به ترتیب اعقاب مزومرها وماکرومرها)محکم به یکدیگر و به لایه ی هیالینی متصل می شوند اما اتصال این سلولها به غشای پایه سست است میکرومها نیز در ابتدا الگوی اتصالی مشابهی را دارند اما الگوی انها حین گاسترو لاسیون تغییر می کند در حالی که سلولهای دیگر اتصال محکم خود را با لایه ی هیالینی و سلولهای مجاور حفظ می کنند پیش سازهای مزانشیمی اولیه تمایل خود را نسبت به این ساختارها از دست می دهند (این تمایل به حدود 2درصد تمایل اولیه می رسد)اما تمایل انها نسبت به ترکیبات موجود درغشا ی پایه و ماده ی زمینه ای برون سلولی (نظیر فیبرونکتین)صد برابر بیشتر می شود
تمامی سلولهای بلاستولا در ابتدا از سطح بیرونی خود به لایه ی هیالینی و از سطح درونی خود به غشای پایه ی ترشح شده توسط خودشان متصل هستند .هر سلول از سطوح جانبی نیز به سلولهای همسایه اتصال دارد.finkوmcclayدریافتند که سلولهای اکتودرمی و اندودرمی اینده ی جنین (به ترتیب اعقاب مزومرها وماکرومرها)محکم به یکدیگر و به لایه ی هیالینی متصل می شوند اما اتصال این سلولها به غشای پایه سست است میکرومها نیز در ابتدا الگوی اتصالی مشابهی را دارند اما الگوی انها حین گاسترو لاسیون تغییر می کند در حالی که سلولهای دیگر اتصال محکم خود را با لایه ی هیالینی و سلولهای مجاور حفظ می کنند پیش سازهای مزانشیمی اولیه تمایل خود را نسبت به این ساختارها از دست می دهند (این تمایل به حدود 2درصد تمایل اولیه می رسد)اما تمایل انها نسبت به ترکیبات موجود درغشا ی پایه و ماده ی زمینه ای برون سلولی (نظیر فیبرونکتین)صد برابر بیشتر می شود
این تغییر در تمایلات سلولی موجب ازادسازی اتصالات سلولی میکرومرها از لایه ی هیالینی خارجی و سلولهای مجاور شده و باعث می شود که از طریق غشای پایه به داخل کشیده شده وبه درون بلاستوسل مهاجرت کنند (شکل a 16-8) این تغییر در تمایلات به دلیل تغییر در مولکولهای سطح سلولی است که در این زمان ایجاد می شودهمچنین پرو تئین هایی نظیر فیبرونکتین اینتگرین لامینین1وکادهرین ها نیز در این مهاجرت سلولی شرکت دارند .
همان گونه که درشکل cوb16-8نشان داده شده غلظت بالایی از مواد برون سلولی در اطراف سلولهای مزانشیمی اولیه ی در حال مهاجرت دیده می شود.در داخل بلاستوسل به نظر می رسید که سلولهای مزانشیمی اولیه در امتداد ماده زمینه ی برون سلولی دیواره بلاستوسل مهاجرت کرده و فیلوپودیای خود رابه سمت جلو گسترده می کنندچندین نوع پروتئین(نظیر فیبرونکتین و نوع خاصی گلیکوپروتئین سولفاته)
در شروع و حفظ این مهاجرت لازم و ضروری هستنداما این علایم جهت یابی کافی نیستند چرا که سلولهای در مهاجرت((میدانند))که چه موقع حرکت خود را متوقف کنند و اسپیکولها را نزدیک استوای بلاستوسل تشكيل دهند.
سلولهای مزانشیم اولیه به صورت یک حلقه در موقعیتی خاص و در امتداد محور جانوری-نباتی سازمان دهی میشوند-بسیاری از سلولهای مزانشیم اولیه در دو ناحیه نزدیک بخش شکمی اینده ی لارو تجمع
یافته با یکدیگر ترکیب شده و شروع به تشکیل اسپیکول می کنند(شکل17-8)
شكل 8-17
اگر میکرومر نشان دار شده ای از یک جنین به بلاستوسل جنین دیگری در مرحله ی گاسترولاسیون تزریق شود به محل صحیح مهاجرت کرده ودر تشکیل اسپیکول های جنینی شرکت می کندتصور می شود که اطلاعات مکانی مورد نیاز توسط سلولهای اکتودرمی اینده وغشا ی پایه ی انه فراهم می شود (شکلa18-8}
تنها سلولهای مزانشیمی اولیه (ونه انواع دیگر سلولها)قادرند به این علایم الگوبندی پاسخ دهند
شكل 8-18
.
millerوهمکارانش در 1995از وجود فیلوپودیا کاملا واضح (با قطر3/0ميكرو متر)در سلولهای مزانشیمی تشکیل دهنده ی اسکلت سلولی خبر دادند
به نظر می رسد که این فیلو پودیا در حرکت نقش فعالی نداشته بلکه به جستجوی دیواره ی بلاستوسل می پردازد وشاید در پاسخگویی به علایم الگو بندی پشتی – شکمی و جانوری نباتی صادر شده از اکتو درم نقش داشته باشد.
مرحله ی اول درون روی آرکنترون
هم زمان که سلول های مزانشیمی اولیه ی قطب نباتی جنین کروی را ترک می کنند ، تغییرات مهمی در سلول های باقی مانده ایجاد می شود.سلول های باقی مانده ضخیم و پهن شده و صفحه ی نباتی را تشکيل دهند.در نتیجه شکل جنین تغییر می کند شكل8 -51)ساعت 9سلول های صفحه ی نباتی به یکدیگر و به لایه ی هیالینی سلول تخم متصل باقی مانده و به گونه ایی حرکت میکنند تا فضاهای خالی به وجود آمده ی حاصل از مهاجرت مزانشیم اولیه را پر کنند. به علاوه صفحه نباتی به درون خم شده وپس از درون روی حدود 4/1الی 2/1به طور ناگهانی متوقف می شوند دهانه این ناحیه درون رفته (آرکنترون ) بلاستو پور نامیده میشود.
بررسیها نشان داده که لایه هیالینی موجوددر صفحه نباتی به دلیل تغییر در ترکیباتش به سمت درون خمیده میشود.لایه هیالینی درواقع از دو تیغه تشکیل شده است یک تیغه بیرونی متشکل از پروتیین هیالین و یک پروتیین درونی متشکل از پروتیین های فیبروپیلین .فیبروپیلین ها در گرانول های ترشحی تخمک ذخیره شده وپس از اگزوسیتوز گرانول های قشری ترشح می شوند و منجر به آزاد سازی پروتیین هیالین می گردند.درمرحله بلاستولا فیبروپلین ها شبکه ای تور ماننددر اطراف جنین ایجاد میکند .در حین درون روی سلول های صفحه نباتی شرو به ترشح گلیکو پروتیین کندرویتین سولفات به درون تیغه درونی لایه هیالینی میکند در نتیجهناحیه نباتی لایه هیالینی خمیده میشود اندکی بعد یک نیروی ثانویه که از حرکت سلولهای اپی تلیالی مجاور صفحه نباتی حاصل میشود با کشیدن لایه خمیده به درون موجب تسهیل در فرآیند درون روی می گردد.
سلول های مزانشیم ثانویه اولین گروه از سلول هایی هستند که درون روی نموده ، به درون بلاستوسل پیش روی می کنند و راس آرکنترون را تشکیل می دهند . آنها نهایتاً سلول های رنگدانه دار ، عضلات اطراف لوله ی گوارشی ، و کیسه های سلومی را تشکیل
می دهند سلول های اندودرمی مجاور مزانشیم مشتق شده از میکرومر ،لوله ی گوارشی قدامی را به وجود می آورند که فاصله ی بیشتری را در بلاستوسل مهاجرت می کند . لایه ی بعد سلول های اندودرمی لوله ی گوارشی میانی را به وجود می آورد . ردیف محیطی آخر نیز درون روی نموده ، لوله ی گوارش خلفی و مخرج را تشکیل می دهد
مراحل دوم و سوم درون روی آرکنترون
دنبال درون روی اول ، فاز دوم شکل گیری آرکنترون شروع می شود . طی این مرحله آرکنترون گسترده شده به گونه ایی که حتی ممکن است طول آن سه برابر شود . در طی این گسترده
شدن لوله ی گوارش کوتاه و عریض به لوله ای باریک و طویل تغییر شکل می دهد . برای این طریق مهاجرت بر روی یکدیگر و پهن شدن باز آرایی می شوند .
پدیده ایی که طی آن سلول ها در هم فرو می روند و موجب نازک شدن بافت شده و هم زمان یه سمت جلو حرکت می کنند بسط همگرا نامیده میشود .
در تمام گو نه های توتیای در یایی مرحله سوم طویل شدن آرکنترون نیز دیده می شود این مرحله انتهایی در اثر نیروی کششی که توسط سلولهای مزانشیم ثانویه ایجاد میشود آغاز میگردد .این سلولها راس آرکنترون را ساخته ودر همان جا مستقر میشوند سلولهای مذکوردر مایع بلاستوسل زواید فیلوپودیایی ایجاد می کنند که به طرف سطح درونی دیواره بلاستوسل کشیده میشود.فیلوپودیاهادرناحیه اتصالی بین بلاستومرهابه دیواره متصل شده سپس با کوتاه شدن خود آرکنترون را به بالا میکشند(شکل های 8-15و 8-21 ساعت 12و13)
. اگرتعدادی از سلول های مزانشیم ثانویه حذف شوند طویل شدن ادمه می یابد، اگرچه سرعت آن کند می شود . در این گونه سلول های مزانشیم ثانویه نقشی اساسی در کشیده شدن آرکنترون به سمت دیوار بلاستوسل حین مرحله ی انتهایی درون روی بازی می کنند. اما آیا فیلوپودیای مزانشیم ثانویه می تواند به هر بخشی از دیواره ی بلاستوسل متصل شود، یا این که هدف خاصی در نیم کره جانوری باید وجود داشته باشد تا اتصال صورت گیرد؟ آیا بخشی از دیواره ی بلاستوسل وجود دارد که متعهد شده تا بخش شکمی لارورابه وجود آورد؟ مطالعات صورت گرفته توسط Hardin وMcCIay در1990 نشان می دهد که ناحیه ی هدف ویژه ای برای فیلوپود یا وجود دارد که از سایر نواحی قطب جانوری متفاوت است فیلوپود یاگسترش می یابد و به صورت تصادفی به دیواره ی بلاستوسل متصل شده وسپس آزاد می گردد. با این وجودزمانی که فیلوپود یا با ناحیه ی خاصی از دیواره برخورد می کند اتصال خود را حفظ کرده ، منقبض شده وآرکنترون را به سمت خود می کشد. زمانی که Hardin وMcCIay سمت دیگر دیواره ی بلاستوسل رادرمعرض فیلوپود یاها قراردادند، فیلوپود یا به گسترش خود ادامه داده وبعد از برخورد با این دیواره ی دست کاری شده آن را رها نمودند. تنها زمانی که فیلوپود یا بافت هدف مورد نظر را پیدا کرد، حرکات خود را متوقف نمود
. اگرگاسترولا به اندازه ای طویل می شد که فیلوپود یا هرگزنمی توانست به ناحیه ی هدف برسد، سلول های مزانشیم ثانویه به جستجوی خود ادمه می دادند وسرانجام آرکنترون را رها کرده وبه صورت آزاد، هدف خود را جستجو می کردند. بنابراین،آنچه واضح است این است که در ناحیه ای که قرار است بخش شکمی لارورا به وجود آورد وهمچنین ناحیه ای از آکنترون که دهان آینده ی جنین را به وجودخواهد آورد،
بخش هدفی وجود دارد که توسط سلول های مزانشیم ثانویه شناسایی می شود . هم زمان با برخورد راُس آرکنترون با ناحیه ی هدف دیواره ی بلا ستوسل سلول های مزانشیم ثانویه روی بلاستوسل پخش می شوند، سپس تکثیر شده واندام های مزودرمی را به وجود می آورند (شکل 15-8 ساعت 5/13). جایی که آرکنترون با دیواربرخورد می کند، نهایتاً به دهان موجود تبدیل خواهد شد
سلول ها مزانشیمی اسکلت زاوارد rudiment شده تا صفحات اسکلتی اولیه سند شوند سمت چپ پلوتئوس سطح دهانی آینده ی جنین توتیای دریایی بالا را به وجود می آورد، حین دگردیسی، imaginal rudime لاروجدا شده سپس لاروتخریب می شود،درحالی که naginal rudiment ( که حالا جوانه نامیده می شود) مجد داً لوله ی گوارش خود ایجاد کرده ودر کف اقیانوس قرار می گیرد. همچنان به مواد غذایی که لاروها شده دریافت کرده وابسته است. الگوی گاسترولاسیون خاربوستان نمونه ی تکاملی تکوین پس دهانیازبه
حساب می آید.درپس دهانیان (خارپوستان ، تونیکات ها سفالوکوردات ها و مهره داران) منفذ اولیه تبدیل به مخرج می شود، درحالیکه منفذ دوم دهان را تشکیل می دهد (دوتروستوم به معنی دهان ثانویه). به علاوه درتوتیای
دریایی پدپده ی گسترش همگرا واستفاده ازبیان ژن Nodal درتثبیت محورها دیده می شود












